

Biochemical differentiation of APOBEC3F and APOBEC3G proteins associated with HIV-1 life cycle
- PMID: 17142455
- PMCID: PMC1880894
- DOI: 10.1074/jbc.M610150200
Biochemical differentiation of APOBEC3F and APOBEC3G proteins associated with HIV-1 life cycle
Abstract
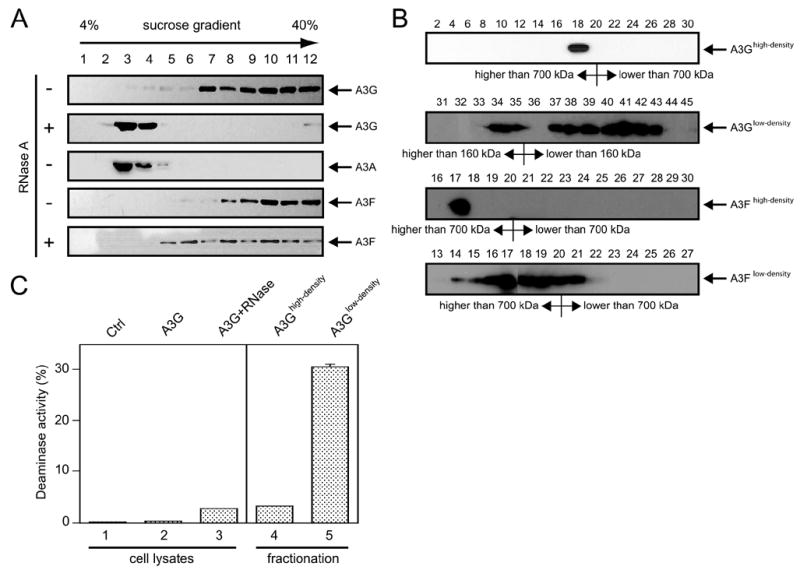
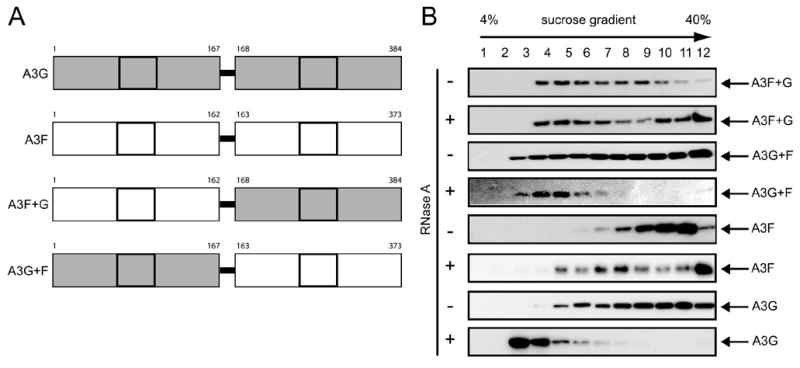
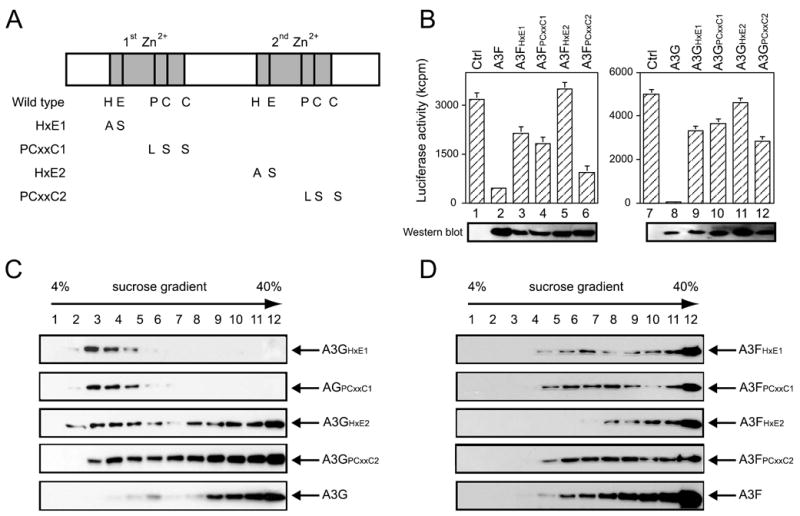
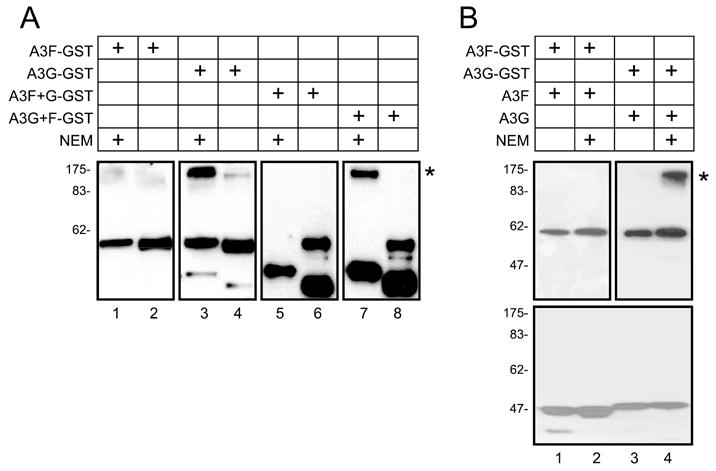
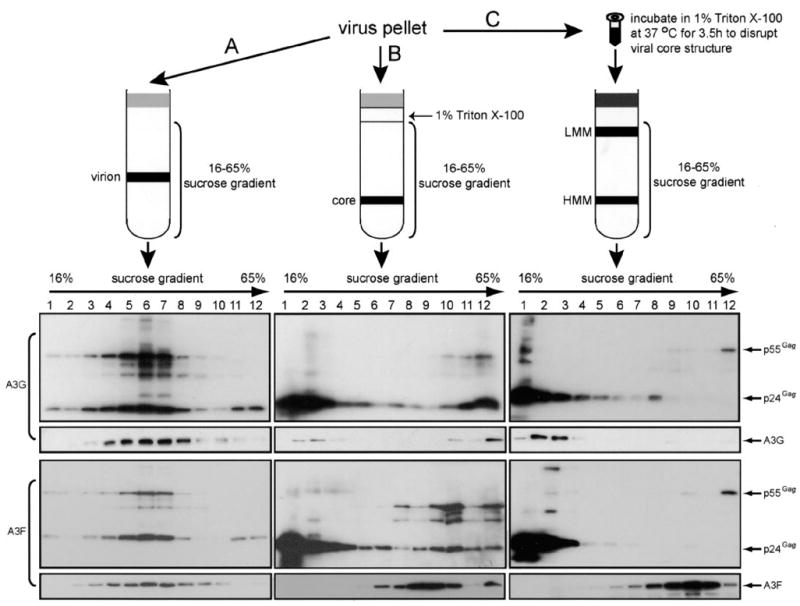
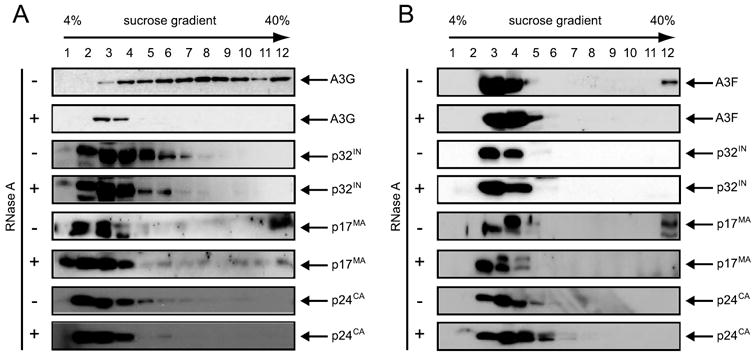
APOBEC3G and APOBEC3F are cytidine deaminase with duplicative cytidine deaminase motifs that restrict HIV-1 replication by catalyzing C-to-U transitions on nascent viral cDNA. Despite 60% protein sequence similarity, APOBEC3F and APOBEC3G have a different target consensus sequence for editing, and importantly, APOBEC3G has 10-fold higher anti-HIV activity than APOBEC3F. Thus, APOBEC3F and APOBEC3G may have distinctive characteristics that account for their functional differences. Here, we have biochemically characterized human APOBEC3F and APOBEC3G protein complexes as a function of the HIV-1 life cycle. APOBEC3G was previously shown to form RNase-sensitive, enzymatically inactive, high molecular mass complexes in immortalized cells, which are converted into enzymatically active, low molecular mass complexes by RNase digestion. We found that APOBEC3F also formed high molecular mass complexes in these cells, but these complexes were resistant to RNase treatment. Further, the N-terminal half determined RNase sensitivity and was necessary for the high molecular mass complex assembly of APOBEC3G but not APOBEC3F. Unlike APOBEC3F, APOBEC3G strongly interacted with cellular proteins via disulfide bonds. Inside virions, both APOBEC3F and APOBEC3G were found in viral cores, but APOBEC3G was associated with low molecular mass, whereas APOBEC3F was still retained in high molecular mass complexes. After cell entry, both APOBEC3F and APOBEC3G were localized in low molecular mass complexes associated with viral reverse transcriptional machinery. These results demonstrate that APOBEC3F and APOBEC3G complexes undergo dynamic conversion during HIV-1 infection and also reveal biochemical differences that likely determine their different anti-HIV-1 activity.
Figures
Similar articles
-
APOBEC3F can inhibit the accumulation of HIV-1 reverse transcription products in the absence of hypermutation. Comparisons with APOBEC3G.J Biol Chem. 2007 Jan 26;282(4):2587-95. doi: 10.1074/jbc.M607298200. Epub 2006 Nov 22. J Biol Chem. 2007. PMID: 17121840
-
Comparative analysis of the antiretroviral activity of APOBEC3G and APOBEC3F from primates.Virology. 2006 May 25;349(1):31-40. doi: 10.1016/j.virol.2005.12.035. Epub 2006 Feb 7. Virology. 2006. PMID: 16460778
-
Mutational comparison of the single-domained APOBEC3C and double-domained APOBEC3F/G anti-retroviral cytidine deaminases provides insight into their DNA target site specificities.Nucleic Acids Res. 2005 Apr 4;33(6):1913-23. doi: 10.1093/nar/gki343. Print 2005. Nucleic Acids Res. 2005. PMID: 15809227 Free PMC article.
-
Antiviral roles of APOBEC proteins against HIV-1 and suppression by Vif.Arch Virol. 2009;154(10):1579-88. doi: 10.1007/s00705-009-0481-y. Epub 2009 Aug 12. Arch Virol. 2009. PMID: 19669862 Review.
-
APOBEC deaminases as cellular antiviral factors: a novel natural host defense mechanism.Med Sci Monit. 2006 May;12(5):RA92-8. Med Sci Monit. 2006. PMID: 16641889 Review.
Cited by
-
The role of cytidine deaminases on innate immune responses against human viral infections.Biomed Res Int. 2013;2013:683095. doi: 10.1155/2013/683095. Epub 2013 Jun 25. Biomed Res Int. 2013. PMID: 23865062 Free PMC article. Review.
-
The in vitro Biochemical Characterization of an HIV-1 Restriction Factor APOBEC3F: Importance of Loop 7 on Both CD1 and CD2 for DNA Binding and Deamination.J Mol Biol. 2016 Jul 3;428(13):2661-70. doi: 10.1016/j.jmb.2016.03.031. Epub 2016 Apr 8. J Mol Biol. 2016. PMID: 27063502 Free PMC article.
-
Positioning of APOBEC3G/F mutational hotspots in the human immunodeficiency virus genome favors reduced recognition by CD8+ T cells.PLoS One. 2014 Apr 10;9(4):e93428. doi: 10.1371/journal.pone.0093428. eCollection 2014. PLoS One. 2014. PMID: 24722422 Free PMC article.
-
Molecular Interactions of a DNA Modifying Enzyme APOBEC3F Catalytic Domain with a Single-Stranded DNA.J Mol Biol. 2018 Jan 5;430(1):87-101. doi: 10.1016/j.jmb.2017.11.007. Epub 2017 Nov 27. J Mol Biol. 2018. PMID: 29191651 Free PMC article.
-
Associations between activation-induced cytidine deaminase/apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like cytidine deaminase expression, hepatitis B virus (HBV) replication and HBV-associated liver disease (Review).Mol Med Rep. 2015 Nov;12(5):6405-14. doi: 10.3892/mmr.2015.4312. Epub 2015 Sep 10. Mol Med Rep. 2015. PMID: 26398702 Free PMC article. Review.
References
-
- Conticello SG, Thomas CJ, Petersen-Mahrt SK, Neuberger MS. Mol Biol Evol. 2005;22:367–377. - PubMed
-
- Jarmuz A, Chester A, Bayliss J, Gisbourne J, Dunham I, Scott J, Navaratnam N. Genomics. 2002;79:285–296. - PubMed
-
- Rogozin IB, Basu MK, Jordan IK, Pavlov YI, Koonin EV. Cell Cycle. 2005;4:1281–1285. - PubMed
-
- Wedekind JE, Dance GS, Sowden MP, Smith HC. Trends Genet. 2003;19:207–216. - PubMed
-
- MacGinnitie AJ, Anant S, Davidson NO. J Biol Chem. 1995;270:14768–14775. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
