

Functional analysis of SPINDLY in gibberellin signaling in Arabidopsis
- PMID: 17142481
- PMCID: PMC1803720
- DOI: 10.1104/pp.106.091025
Functional analysis of SPINDLY in gibberellin signaling in Arabidopsis
Abstract
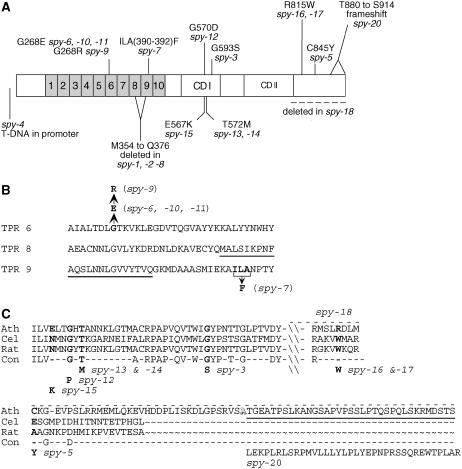
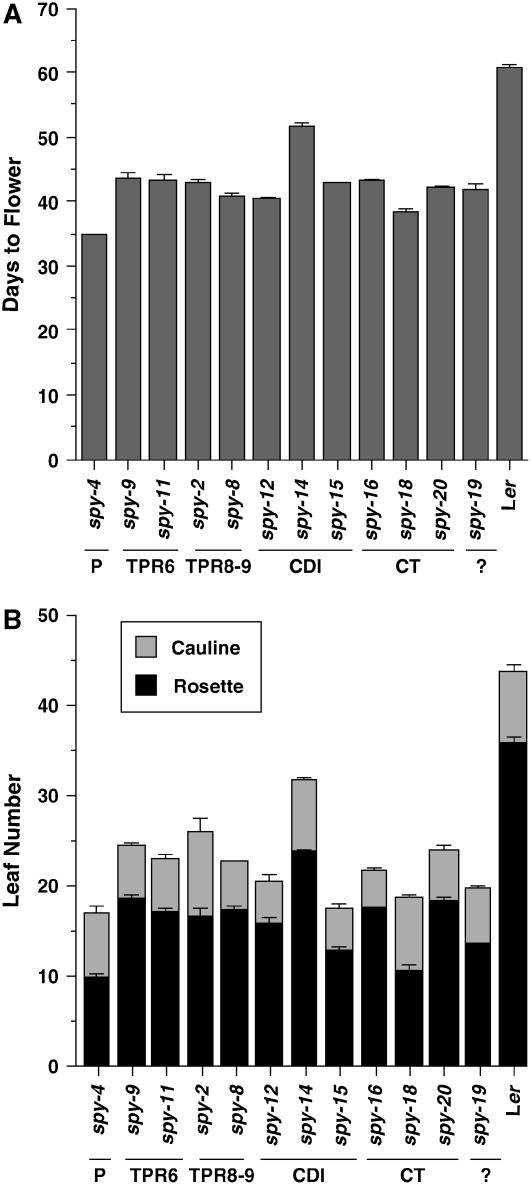
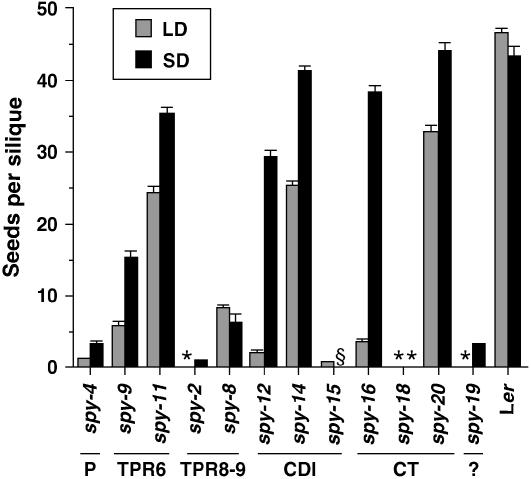
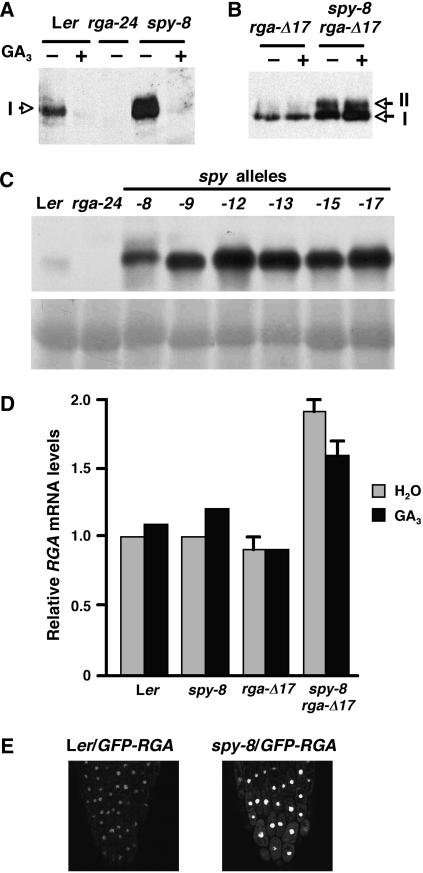
The Arabidopsis (Arabidopsis thaliana) SPINDLY (SPY) protein negatively regulates the gibberellin (GA) signaling pathway. SPY is an O-linked N-acetylglucosamine (GlcNAc) transferase (OGT) with a protein-protein interaction domain consisting of 10 tetratricopeptide repeats (TPR). OGTs add a GlcNAc monosaccharide to serine/threonine residues of nuclear and cytosolic proteins. Determination of the molecular defects in 14 new spy alleles reveals that these mutations cluster in three TPRs and the C-terminal catalytic region. Phenotypic characterization of 12 spy alleles indicates that TPRs 6, 8, and 9 and the catalytic domain are crucial for GA-regulated stem elongation, floral induction, and fertility. TPRs 8 and 9 and the catalytic region are also important for modulating trichome morphology and inflorescence phyllotaxy. Consistent with a role for SPY in embryo development, several alleles affect seedling cotyledon number. These results suggest that three of the TPRs and the OGT activity in SPY are required for its function in GA signal transduction. We also examined the effect of spy mutations on another negative regulator of GA signaling, REPRESSOR OF ga1-3 (RGA). The DELLA motif in RGA is essential for GA-induced proteolysis of RGA, and deletion of this motif (as in rga-delta17) causes a GA-insensitive dwarf phenotype. Here, we demonstrate that spy partially suppresses the rga-delta17 phenotype but does not reduce rga-delta17 or RGA protein levels or alter RGA nuclear localization. We propose that SPY may function as a negative regulator of GA response by increasing the activity of RGA, and presumably other DELLA proteins, by GlcNAc modification.
Figures


References
-
- Bailey AL (1995) Single-stranded conformational polymorphisms. In MA Innis, DH Gelfand, JJ Sninsky, eds, PCR Strategies. Academic Press, San Diego, pp 121–129
-
- Blatch GL, Lässle M (1999) The tetratricopeptide repeat: a structural motif mediating protein-protein interactions. Bioessays 21 932–939 - PubMed
-
- Carol P, Peng J, Harberd NP (1995) Isolation and preliminary characterization of gas1-1, a mutation causing partial suppression of the phenotype conferred by the gibberellin-insensitive (gai) mutation in Arabidopsis thaliana (L.) Heynh. Planta 197 414–417 - PubMed
-
- Cheng H, Qin L, Lee S, Fu X, Richards DE, Cao D, Luo D, Harberd NP, Peng J (2004) Gibberellin regulates Arabidopsis floral development via suppression of DELLA protein function. Development 131 1055–1064 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
