

Discrimination in the dark. Resolving the interplay between metabolic and physical constraints to phosphoenolpyruvate carboxylase activity during the crassulacean acid metabolism cycle
- PMID: 17142488
- PMCID: PMC1803711
- DOI: 10.1104/pp.106.088302
Discrimination in the dark. Resolving the interplay between metabolic and physical constraints to phosphoenolpyruvate carboxylase activity during the crassulacean acid metabolism cycle
Abstract
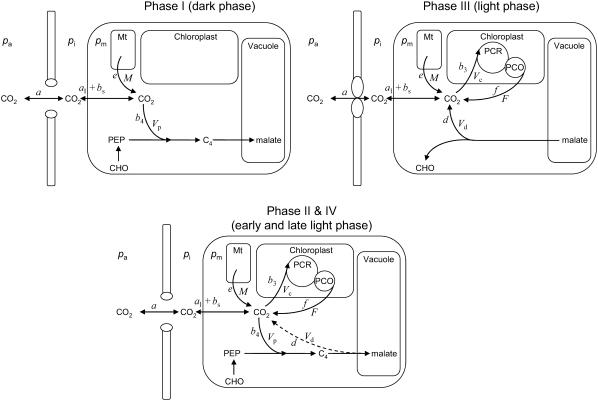
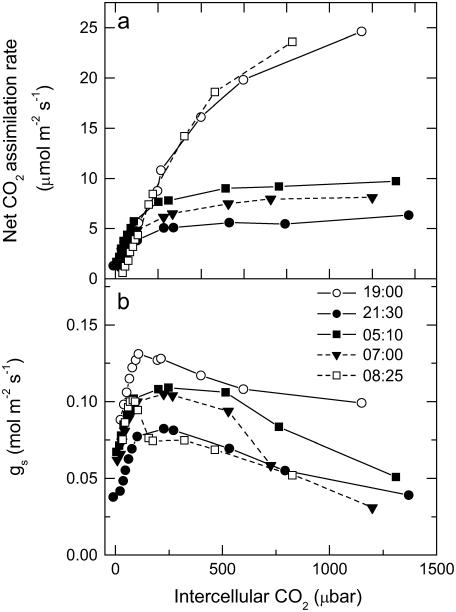
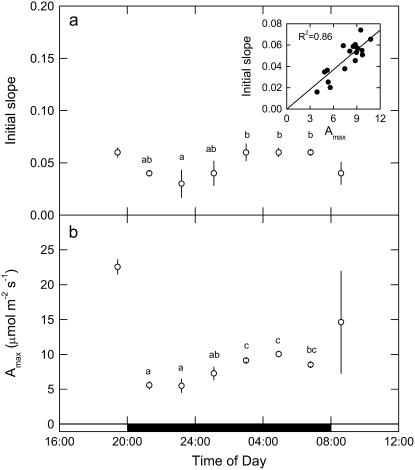
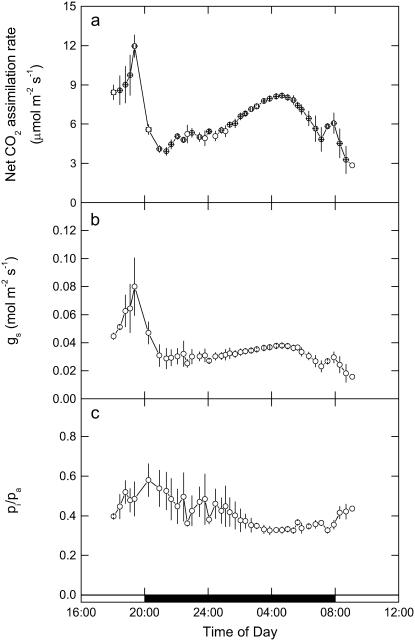
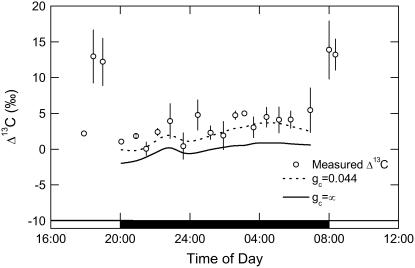
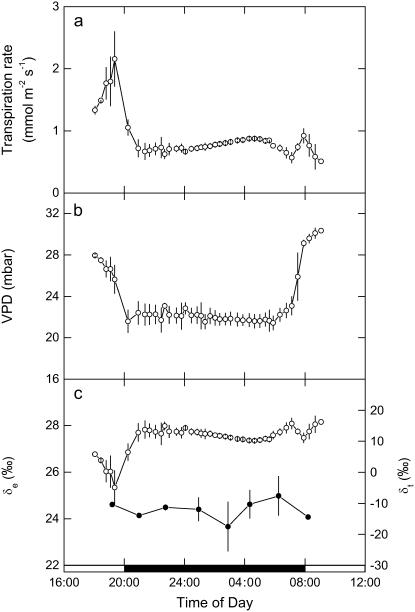
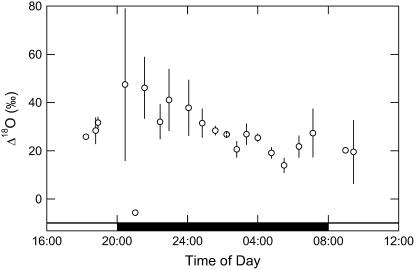
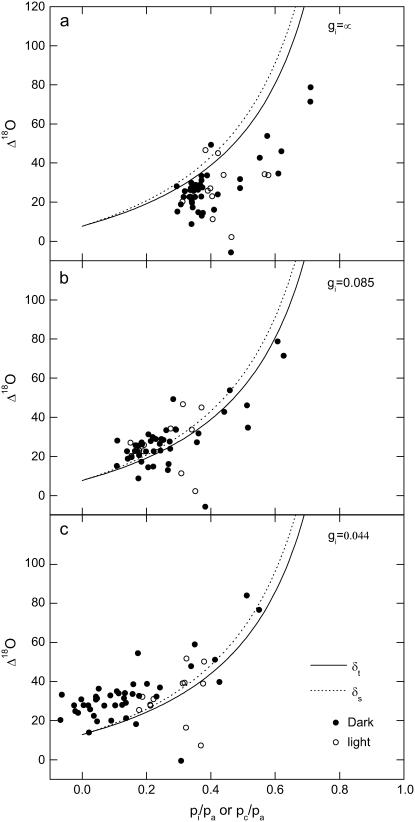
A model defining carbon isotope discrimination (delta13C) for crassulacean acid metabolism (CAM) plants was experimentally validated using Kalanchoe daigremontiana. Simultaneous measurements of gas exchange and instantaneous CO2 discrimination (for 13C and 18O) were made from late photoperiod (phase IV of CAM), throughout the dark period (phase I), and into the light (phase II). Measurements of CO2 response curves throughout the dark period revealed changing phosphoenolpyruvate carboxylase (PEPC) capacity. These systematic changes in PEPC capacity were tracked by net CO2 uptake, stomatal conductance, and online delta13C signal; all declined at the start of the dark period, then increased to a maximum 2 h before dawn. Measurements of delta13C were higher than predicted from the ratio of intercellular to external CO2 (p(i)/p(a)) and fractionation associated with CO2 hydration and PEPC carboxylations alone, such that the dark period mesophyll conductance, g(i), was 0.044 mol m(-2) s(-1) bar(-1). A higher estimate of g(i) (0.085 mol m(-2) s(-1) bar(-1)) was needed to account for the modeled and measured delta18O discrimination throughout the dark period. The differences in estimates of g(i) from the two isotope measurements, and an offset of -5.5 per thousand between the 18O content of source and transpired water, suggest spatial variations in either CO2 diffusion path length and/or carbonic anhydrase activity, either within individual cells or across a succulent leaf. Our measurements support the model predictions to show that internal CO2 diffusion limitations within CAM leaves increase delta13C discrimination during nighttime CO2 fixation while reducing delta13C during phase IV. When evaluating the phylogenetic distribution of CAM, carbon isotope composition will reflect these diffusive limitations as well as relative contributions from C3 and C4 biochemistry.
Figures
Similar articles
-
Contribution of carbon fixed by Rubisco and PEPC to phloem export in the Crassulacean acid metabolism plant Kalanchoe daigremontiana.J Exp Bot. 2010 Mar;61(5):1375-83. doi: 10.1093/jxb/erq006. Epub 2010 Feb 16. J Exp Bot. 2010. PMID: 20159885 Free PMC article.
-
Leaf succulence determines the interplay between carboxylase systems and light use during Crassulacean acid metabolism in Kalanchöe species.J Exp Bot. 2008;59(7):1851-61. doi: 10.1093/jxb/ern085. Epub 2008 Apr 11. J Exp Bot. 2008. PMID: 18408219
-
Short-term changes in carbon-isotope discrimination in the C3-CAM intermediate Clusia minor L. growing in Trinidad.Oecologia. 1993 Sep;95(3):444-453. doi: 10.1007/BF00321001. Oecologia. 1993. PMID: 28314023
-
The efficiency of C(4) photosynthesis under low light conditions: assumptions and calculations with CO(2) isotope discrimination.J Exp Bot. 2011 May;62(9):3119-34. doi: 10.1093/jxb/err073. Epub 2011 Apr 28. J Exp Bot. 2011. PMID: 21527629 Review.
-
Achievable productivities of certain CAM plants: basis for high values compared with C3 and C4 plants.New Phytol. 1991 Oct;119(2):183-205. doi: 10.1111/j.1469-8137.1991.tb01022.x. New Phytol. 1991. PMID: 33874131 Review.
Cited by
-
Transgenic maize phosphoenolpyruvate carboxylase alters leaf-atmosphere CO2 and 13CO2 exchanges in Oryza sativa.Photosynth Res. 2019 Nov;142(2):153-167. doi: 10.1007/s11120-019-00655-4. Epub 2019 Jul 19. Photosynth Res. 2019. PMID: 31325077 Free PMC article.
-
Effect of Light/Dark Cycle on Photosynthetic Pathway Switching and CO2 Absorption in Two Dendrobium Species.Front Plant Sci. 2019 May 22;10:659. doi: 10.3389/fpls.2019.00659. eCollection 2019. Front Plant Sci. 2019. PMID: 31178881 Free PMC article.
-
The role of cis-elements in the evolution of crassulacean acid metabolism photosynthesis.Hortic Res. 2020 Jan 1;7:5. doi: 10.1038/s41438-019-0229-0. eCollection 2020. Hortic Res. 2020. PMID: 31908808 Free PMC article. Review.
-
Crassulacean acid metabolism-cycling in Euphorbia milii.AoB Plants. 2013;5:plt014. doi: 10.1093/aobpla/plt014. Epub 2013 Apr 17. AoB Plants. 2013. PMID: 23596548 Free PMC article.
-
The evolution of inorganic carbon concentrating mechanisms in photosynthesis.Philos Trans R Soc Lond B Biol Sci. 2008 Aug 27;363(1504):2641-50. doi: 10.1098/rstb.2008.0020. Philos Trans R Soc Lond B Biol Sci. 2008. PMID: 18487130 Free PMC article. Review.
References
-
- Borland AM, Dodd AN (2002) Carbohydrate partitioning in crassulacean acid metabolism plants: reconciling potential conflicts of interest. Funct Plant Biol 29 707–716 - PubMed
-
- Borland AM, Griffiths H (1997) A comparative study on the regulation of C-3 and C-4 carboxylation processes in the constitutive crassulacean acid metabolism (CAM) plant Kalanchoe daigremontiana and the C-3-CAM intermediate Clusia minor. Planta 201 368–378 - PubMed
-
- Borland AM, Griffiths H, Broadmeadow MSJ, Fordham MC, Maxwell C (1993) Short-term changes in carbon-isotope discrimination in the C3-CAM intermediate Clusia minor L growing in Trinidad. Oecologia 95 444–453 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
