

Th2 cell-selective enhancement of human IL13 transcription by IL13-1112C>T, a polymorphism associated with allergic inflammation
- PMID: 17142763
- PMCID: PMC11507172
- DOI: 10.4049/jimmunol.177.12.8633
Th2 cell-selective enhancement of human IL13 transcription by IL13-1112C>T, a polymorphism associated with allergic inflammation
Abstract
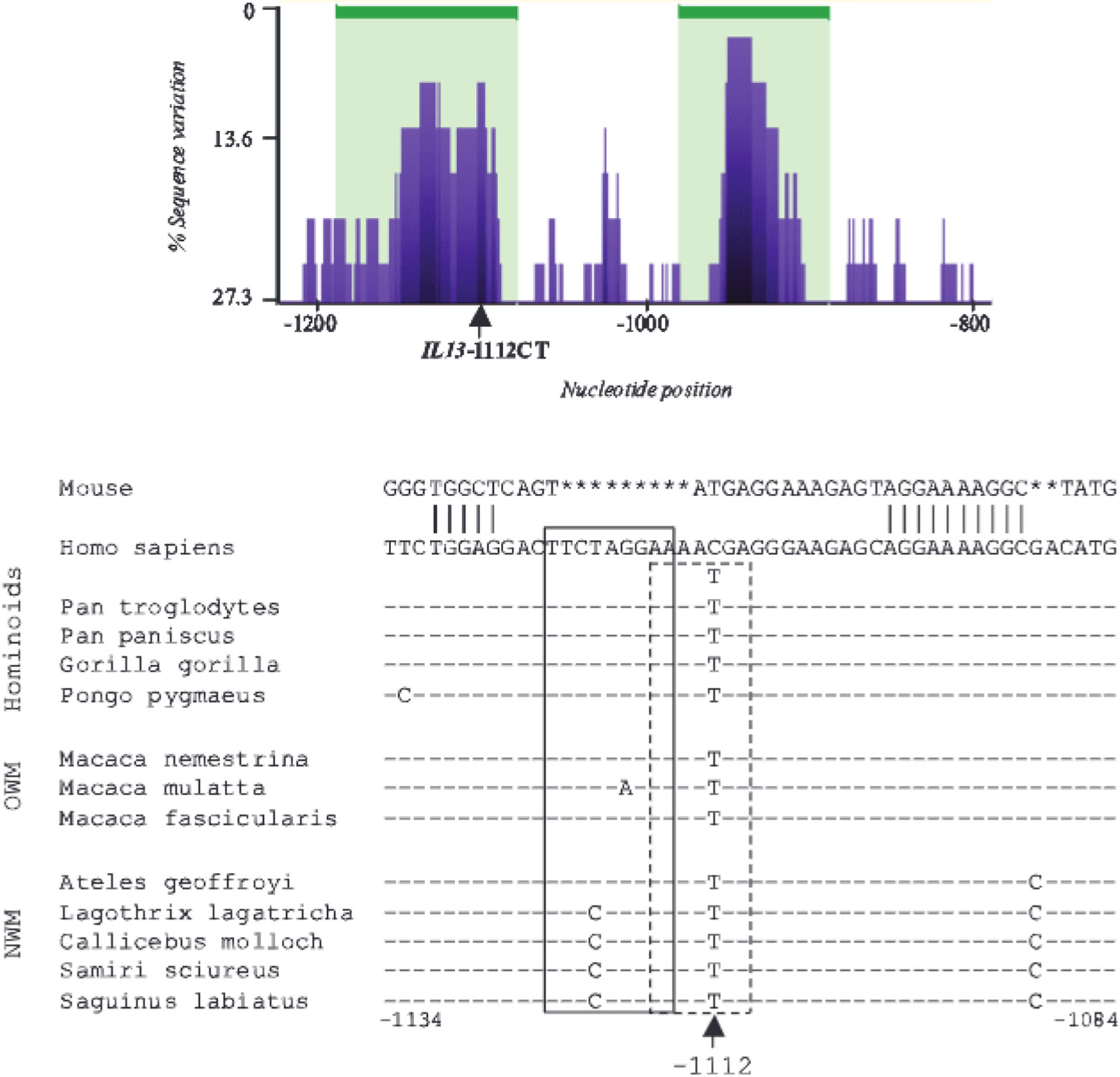
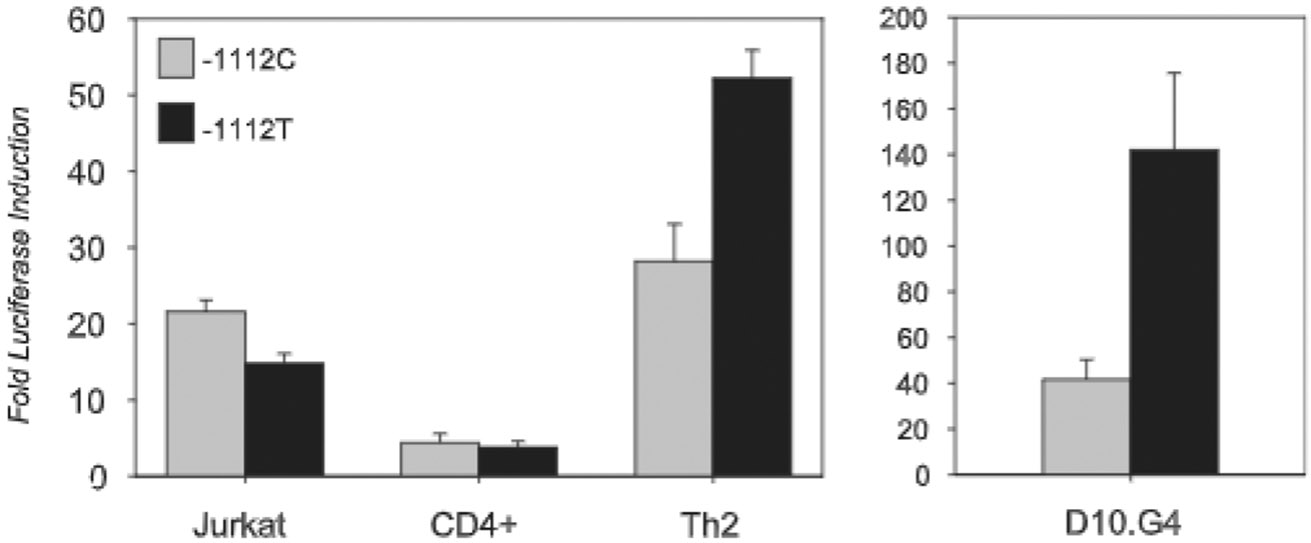
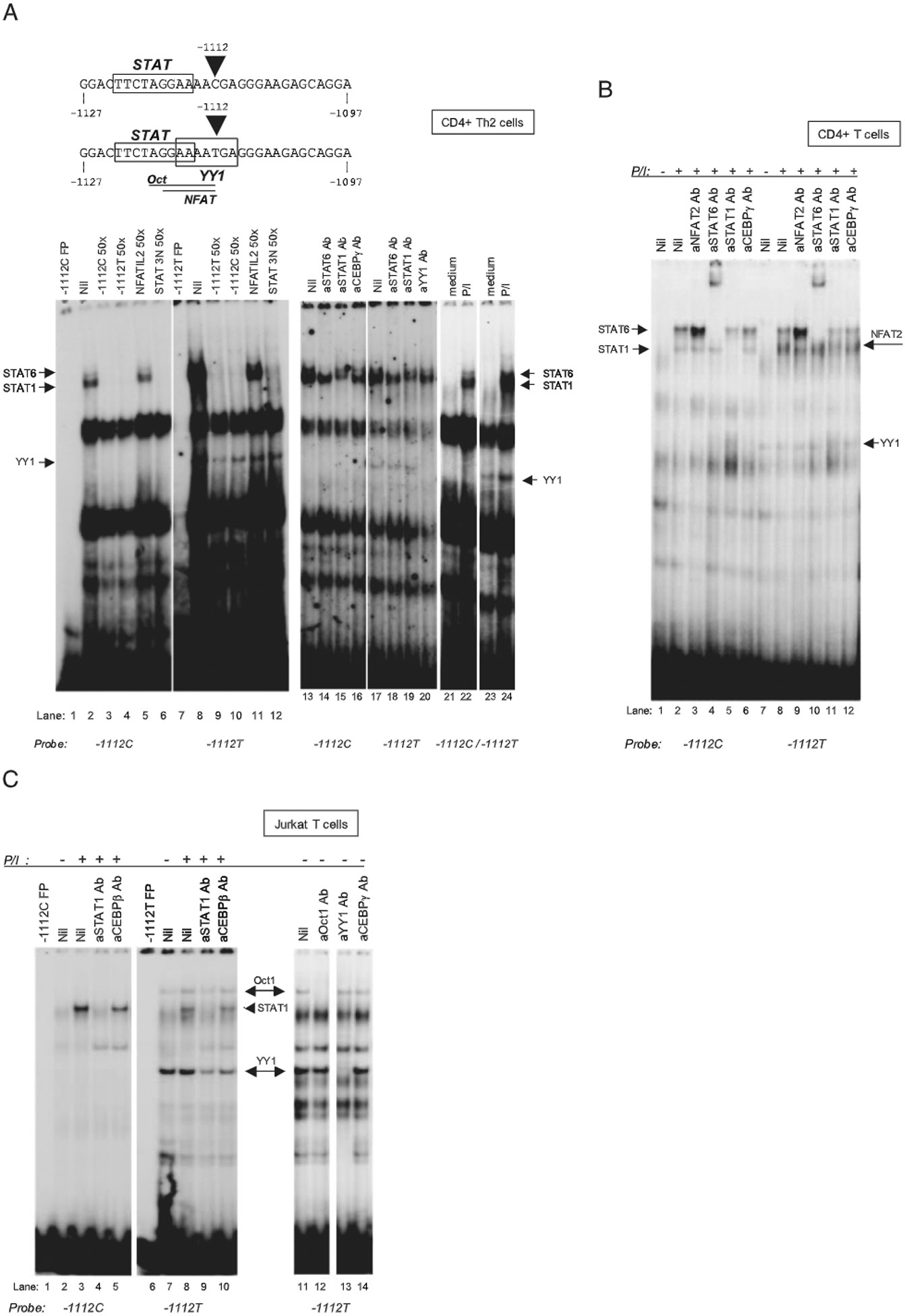
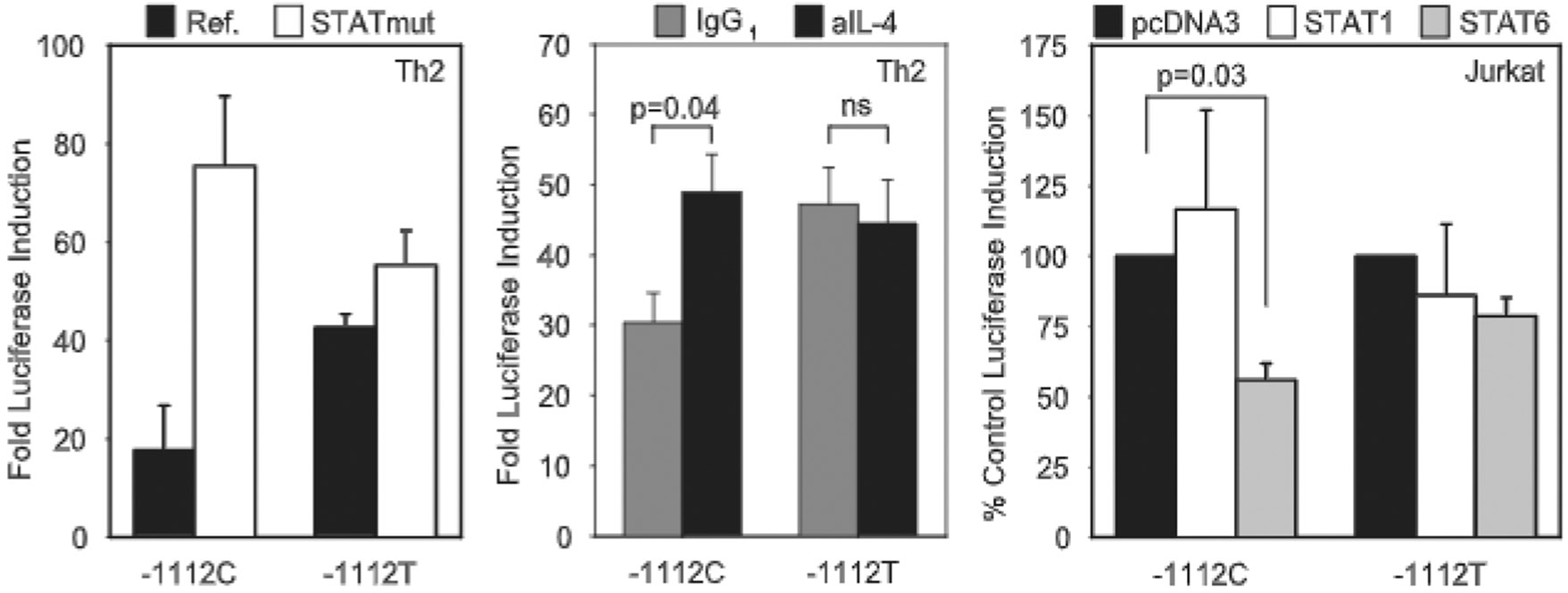
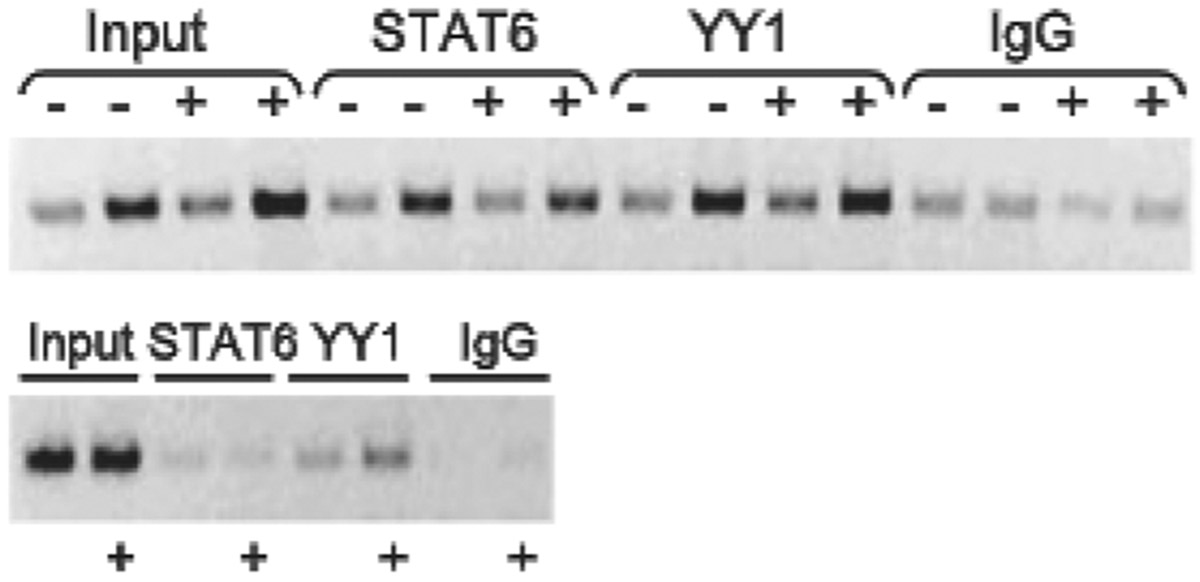
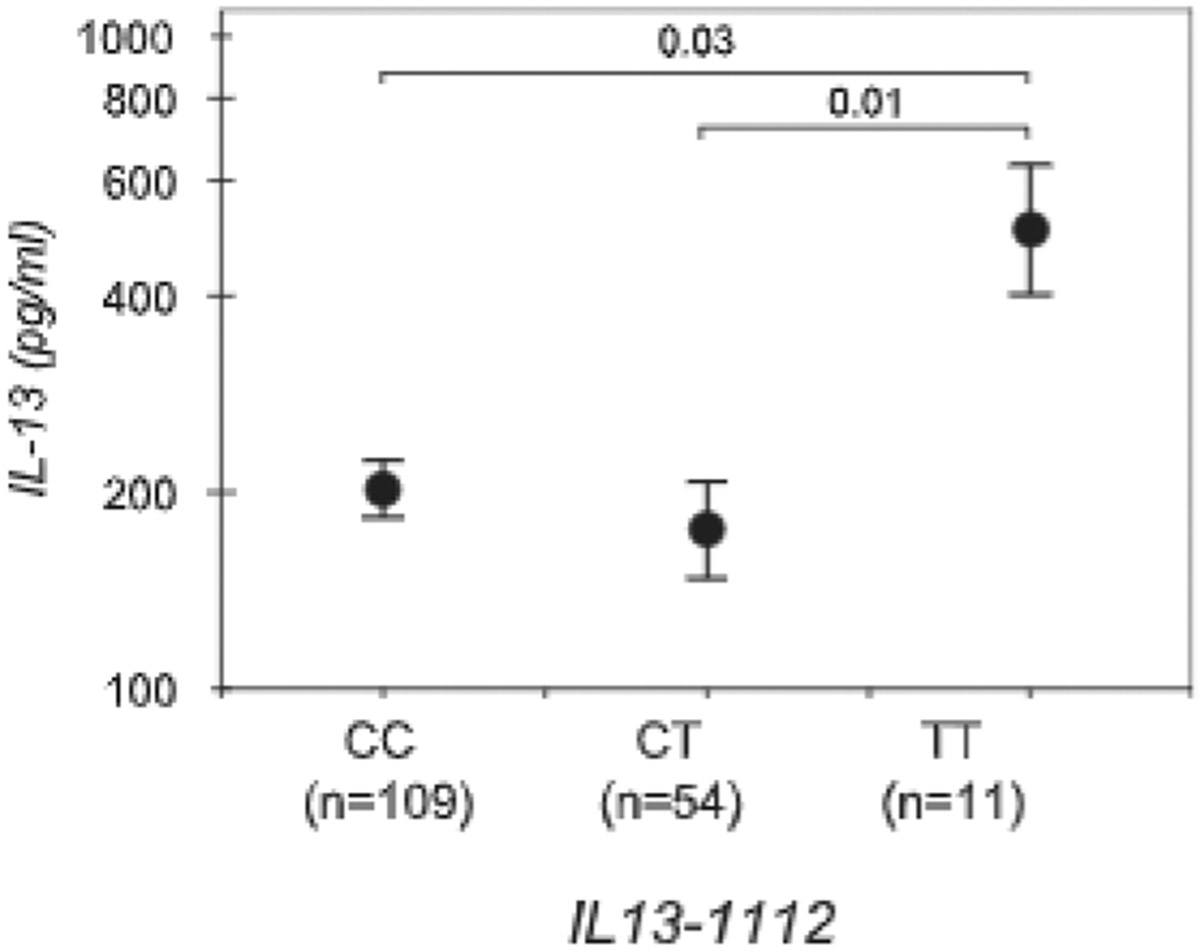
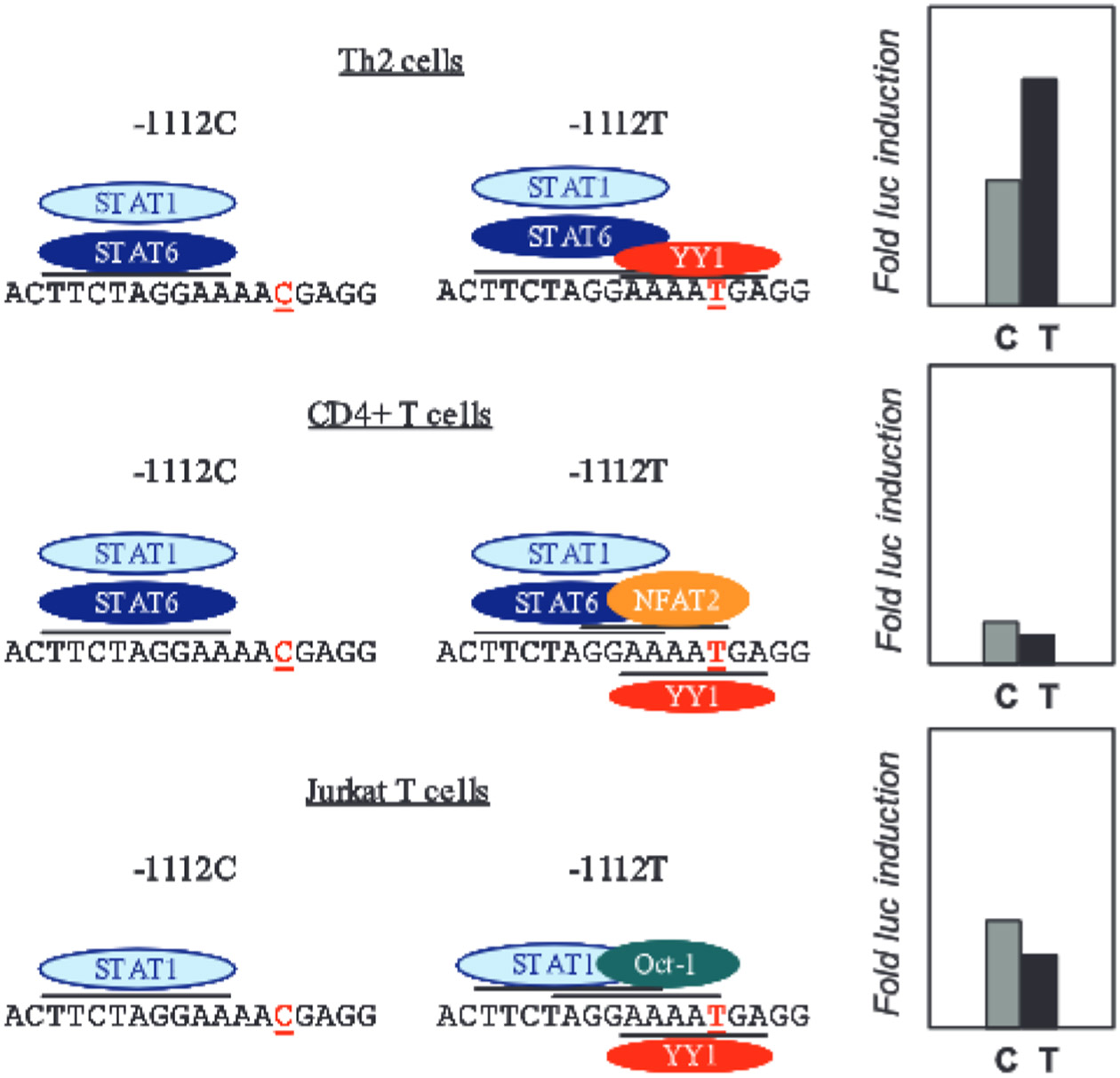
IL-13 is a central mediator of allergic inflammation. The single nucleotide polymorphism IL13-1112C>T (rs1800925) is associated with allergic phenotypes in ethnically distinct populations, but the underlying mechanism(s) remain unknown. Using in vivo, in vitro, and in silico analysis, we show that the IL13-1112T allele enhanced IL13 promoter activity in primary human and murine CD4(+) Th2 lymphocytes. Increased expression of IL13-1112T in Th2 cells was associated with the creation of a Yin-Yang 1 binding site that overlapped a STAT motif involved in negative regulation of IL13 expression and attenuated STAT6-mediated transcriptional repression. Because IL-13 secretion was increased in IL13-1112TT homozygotes, we propose that increased expression of IL13-1112T in vivo may underlie its association with susceptibility to allergic inflammation. Interestingly, IL13-1112T had opposite transcriptional effects in nonpolarized CD4(+) T cells, paralleled by distinct patterns of DNA-protein interactions at the IL13 promoter. Our findings suggest the nuclear milieu dictates the functional outcome of genetic variation.
Conflict of interest statement
Disclosures
The authors have no financial conflict of interest.
Figures
Similar articles
-
An allergy-associated polymorphism in a novel regulatory element enhances IL13 expression.Hum Mol Genet. 2009 Dec 1;18(23):4513-20. doi: 10.1093/hmg/ddp411. Epub 2009 Aug 25. Hum Mol Genet. 2009. PMID: 19706623 Free PMC article.
-
NF45 and NF90 regulate HS4-dependent interleukin-13 transcription in T cells.J Biol Chem. 2010 Mar 12;285(11):8256-67. doi: 10.1074/jbc.M109.041004. Epub 2010 Jan 5. J Biol Chem. 2010. PMID: 20051514 Free PMC article.
-
IL-13 R130Q, a common variant associated with allergy and asthma, enhances effector mechanisms essential for human allergic inflammation.J Clin Invest. 2005 Mar;115(3):747-54. doi: 10.1172/JCI22818. J Clin Invest. 2005. PMID: 15711639 Free PMC article.
-
IL13 promoter polymorphism 1112C/T modulates the adverse effect of tobacco smoking on lung function.Am J Respir Crit Care Med. 2007 Oct 15;176(8):748-52. doi: 10.1164/rccm.200704-543OC. Epub 2007 Jul 5. Am J Respir Crit Care Med. 2007. PMID: 17615386
-
Chromatin landscape dynamics of the Il4-Il13 locus during T helper 1 and 2 development.Proc Natl Acad Sci U S A. 2004 Aug 3;101(31):11410-5. doi: 10.1073/pnas.0403334101. Epub 2004 Jul 22. Proc Natl Acad Sci U S A. 2004. PMID: 15272080 Free PMC article.
Cited by
-
Multiple sclerosis susceptibility may be associated with the coding rs20541 (R130Q) IL-13 gene polymorphism in the Polish population.Sci Rep. 2023 Dec 12;13(1):22083. doi: 10.1038/s41598-023-49615-y. Sci Rep. 2023. PMID: 38086930 Free PMC article.
-
Genetic susceptibility to allergic bronchopulmonary aspergillosis in asthma: a genetic association study.Allergy Asthma Clin Immunol. 2016 Sep 27;12:47. doi: 10.1186/s13223-016-0152-y. eCollection 2016. Allergy Asthma Clin Immunol. 2016. PMID: 27708669 Free PMC article.
-
Functional polymorphisms in IL13 are protective against high Schistosoma mansoni infection intensity in a Brazilian population.PLoS One. 2012;7(5):e35863. doi: 10.1371/journal.pone.0035863. Epub 2012 May 4. PLoS One. 2012. PMID: 22574126 Free PMC article.
-
An allergy-associated polymorphism in a novel regulatory element enhances IL13 expression.Hum Mol Genet. 2009 Dec 1;18(23):4513-20. doi: 10.1093/hmg/ddp411. Epub 2009 Aug 25. Hum Mol Genet. 2009. PMID: 19706623 Free PMC article.
-
IL13 gene polymorphisms modify the effect of exposure to tobacco smoke on persistent wheeze and asthma in childhood, a longitudinal study.Respir Res. 2008 Jan 10;9(1):2. doi: 10.1186/1465-9921-9-2. Respir Res. 2008. PMID: 18186920 Free PMC article.
References
-
- Obcr C, and Hoffjan S. 2006. Asthma genetics 2006: the long and winding road to gene discovery. Genes Immun. 7: 95–100. - PubMed
-
- Wills-Karp M, Luyimbazi J, Xu X, Schofield B, Neben TY, Karp CL, and Donaldson DD. 1998. Interleukin-13: central mediator of allergic asthma. Science 282: 2258–2261. - PubMed
-
- Ghaffar O, Laberge S, Jacobson MR, Lowhagen O, Rak S, Durham SR, and Hamid Q. 1997. IL-13 mRNA and immunoreactivity in allergen-induced rhinitis: comparison with IL-4 expression and modulation by topical glucocorticoid therapy. Am. J. Respir. Cell. Mol. Biol 17: 17–24. - PubMed
-
- Lordan JL, Bucchieri F, Richter A, Konstantinidis A, Holloway JW, Thornber M, Puddicombe SM, Buchanan D, Wilson SJ, Djukanovic R, Holgate ST, and Davies DE. 2002. Cooperative effects of Th2 cytokines and allergen on normal and asthmatic bronchial epithelial cells. J. Immunol 169: 407–414. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
