

In vivo administration of D609 leads to protection of subsequently isolated gerbil brain mitochondria subjected to in vitro oxidative stress induced by amyloid beta-peptide and other oxidative stressors: relevance to Alzheimer's disease and other oxidative stress-related neurodegenerative disorders
- PMID: 17145558
- PMCID: PMC1827074
- DOI: 10.1016/j.freeradbiomed.2006.09.002
In vivo administration of D609 leads to protection of subsequently isolated gerbil brain mitochondria subjected to in vitro oxidative stress induced by amyloid beta-peptide and other oxidative stressors: relevance to Alzheimer's disease and other oxidative stress-related neurodegenerative disorders
Abstract
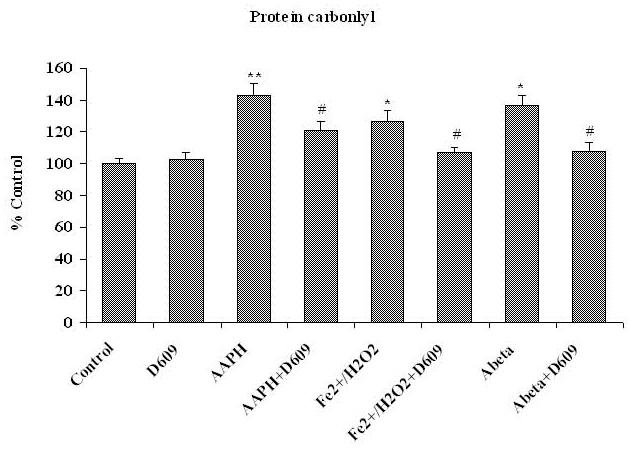
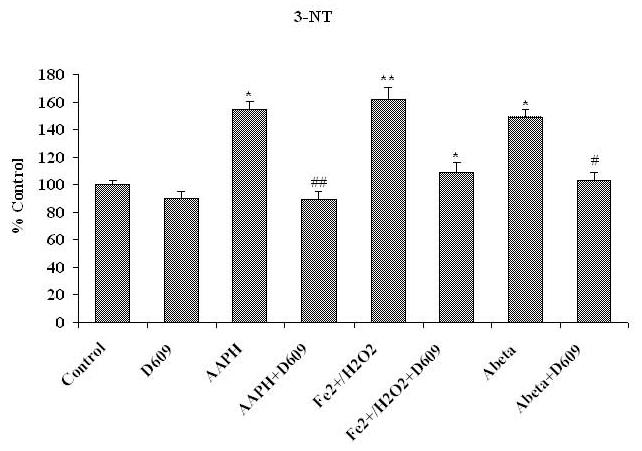
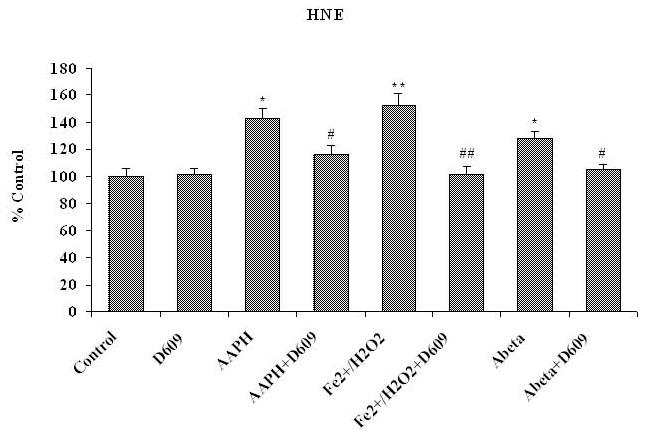
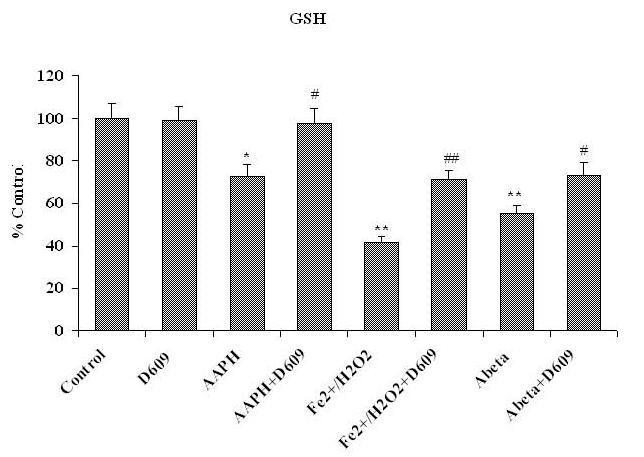
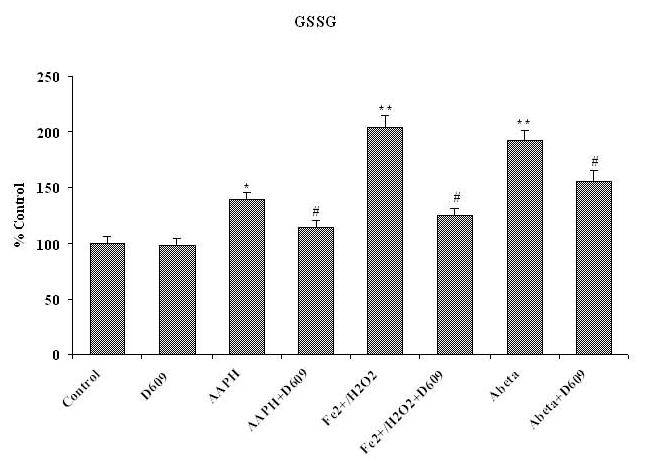
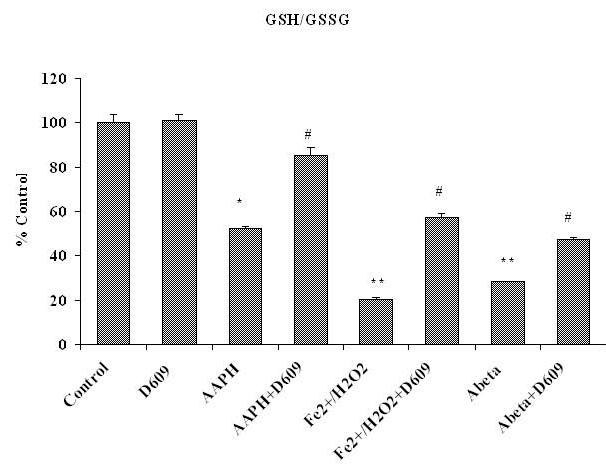
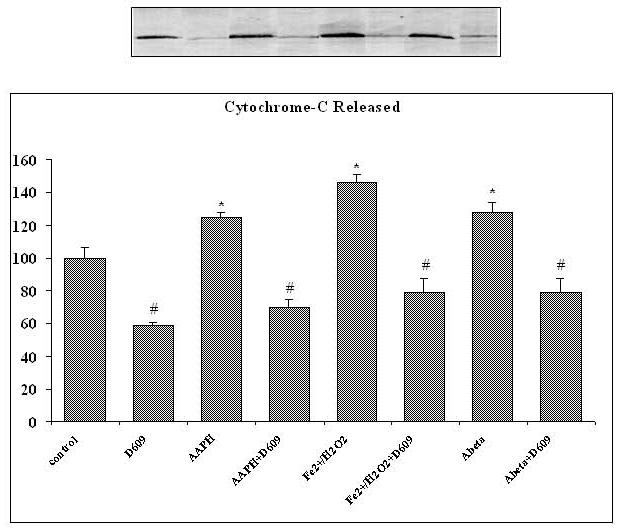
Tricyclodecan-9-yl-xanthogenate (D609) has in vivo and in vitro antioxidant properties. D609 mimics glutathione (GSH) and has a free thiol group, which upon oxidation forms a disulfide. The resulting dixanthate is a substrate for glutathione reductase, regenerating D609. Recent studies have also shown that D609 protects brain in vivo and neuronal cultures in vitro against the potential Alzheimer's disease (AD) causative factor, Abeta(1-42)-induced oxidative stress and cytotoxicity. Mitochondria are important organelles with both pro- and antiapoptotic factor proteins. The present study was undertaken to test the hypothesis that intraperitoneal injection of D609 would provide neuroprotection against free radical-induced, mitochondria-mediated apoptosis in vitro. Brain mitochondria were isolated from gerbils 1 h post injection intraperitoneally (ip) with D609 and subsequently treated in vitro with the oxidants Fe(2+)/H(2)O(2) (hydroxyl free radicals), 2,2-azobis-(2-amidinopropane) dihydrochloride (AAPH, alkoxyl and peroxyl free radicals), and AD-relevant amyloid beta-peptide 1-42 [Abeta(1-42)]. Brain mitochondria isolated from the gerbils previously injected ip with D609 and subjected to these oxidative stress inducers, in vitro, showed significant reduction in levels of protein carbonyls, protein-bound hydroxynonenal [a lipid peroxidation product], 3-nitrotyrosine, and cytochrome c release compared to oxidant-treated brain mitochondria isolated from saline-injected gerbils. D609 treatment significantly maintains the GSH/GSSG ratio in oxidant-treated mitochondria. Increased activity of glutathione S-transferase, glutathione peroxidase, and glutathione reductase in brain isolated from D609-injected gerbils is consistent with the notion that D609 acts like GSH. These antiapoptotic findings are discussed with reference to the potential use of this brain-accessible glutathione mimetic in the treatment of oxidative stress-related neurodegenerative disorders, including AD.
Figures
References
-
- Hensley K, Hall N, Subramaniam R, Cole P, Harris M, Aksenov M, Aksenova M, Gabbita SP, Wu JF, Carney JM, et al. Brain regional correspondence between Alzheimer’s disease histopathology and biomarkers of protein oxidation. J Neurochem. 1995;65::2146–2156. - PubMed
-
- Markesbery WR, Lovell MA. Four-hydroxynonenal, a product of lipid peroxidation, is increased in the brain in Alzheimer’s disease. Neurobiol Aging. 1998;19::33–36. - PubMed
-
- Butterfield DA, Lauderback CM. Lipid peroxidation and protein oxidation in Alzheimer’s disease brain: potential causes and consequences involving amyloid beta-peptide-associated free radical oxidative stress. Free Radic Biol Med. 2002;32::1050–1060. - PubMed
-
- Butterfield DA, Drake J, Pocernich C, Castegna A. Evidence of oxidative damage in Alzheimer’s disease brain: central role for amyloid beta-peptide. Trends Mol Med. 2001;7::548–554. - PubMed
-
- Bosetti F, Brizzi F, Barogi S, Mancuso M, Siciliano G, Tendi EA, Murri L, Rapoport SI, Solaini G. Cytochrome c oxidase and mitochondrial F1F0-ATPase (ATP synthase) activities in platelets and brain from patients with Alzheimer’s disease. Neurobiol Aging. 2002;23::371–376. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
