

Inhibition of BACH1 (FANCJ) helicase by backbone discontinuity is overcome by increased motor ATPase or length of loading strand
- PMID: 17145708
- PMCID: PMC1751539
- DOI: 10.1093/nar/gkl964
Inhibition of BACH1 (FANCJ) helicase by backbone discontinuity is overcome by increased motor ATPase or length of loading strand
Abstract
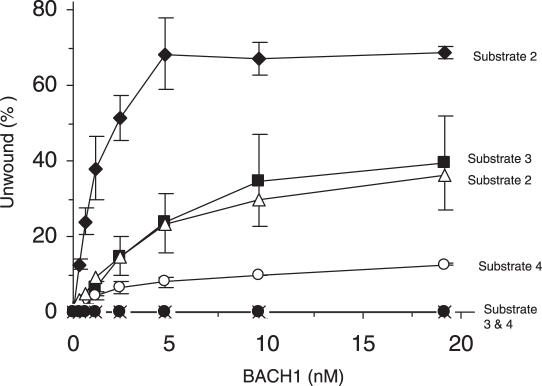
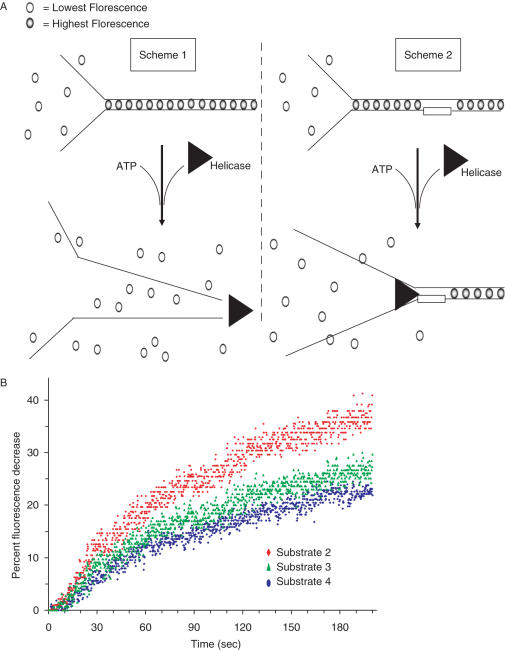
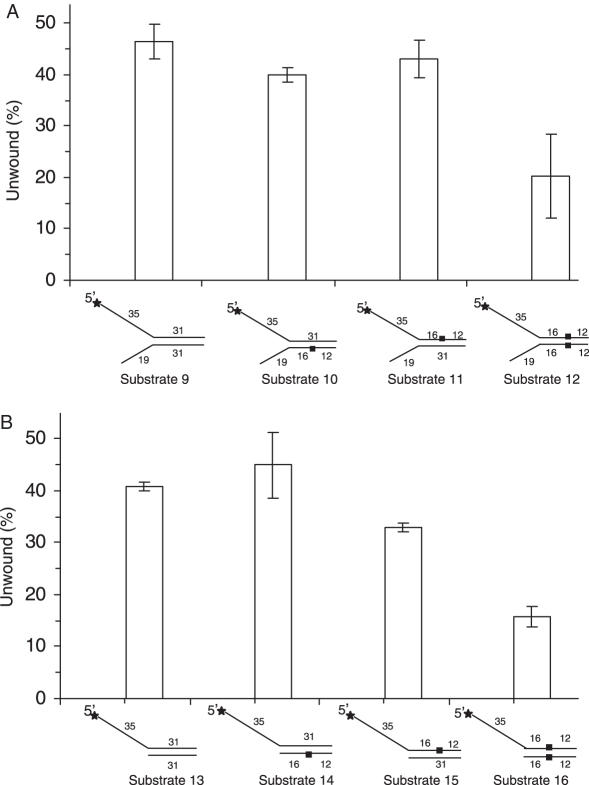
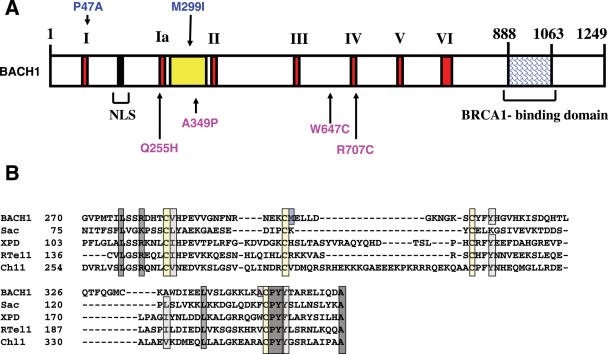
The BRCA1 associated C-terminal helicase (BACH1) associated with breast cancer has been implicated in double strand break (DSB) repair. More recently, BACH1 (FANCJ) has been genetically linked to the chromosomal instability disorder Fanconi Anemia (FA). Understanding the roles of BACH1 in cellular DNA metabolism and how BACH1 dysfunction leads to tumorigenesis requires a comprehensive investigation of its catalytic mechanism and molecular functions in DNA repair. In this study, we have determined that BACH1 helicase contacts with both the translocating and the non-translocating strands of the duplex are critical for its ability to track along the sugar phosphate backbone and unwind dsDNA. An increased motor ATPase of a BACH1 helicase domain variant (M299I) enabled the helicase to unwind the backbone-modified DNA substrate in a more proficient manner. Alternatively, increasing the length of the 5' tail of the DNA substrate allowed BACH1 to overcome the backbone discontinuity, suggesting that BACH1 loading mechanism is critical for its ability to unwind damaged DNA molecules.
Figures



References
-
- Cantor S.B., Bell D.W., Ganesan S., Kass E.M., Drapkin R., Grossman S., Wahrer D.C., Sgroi D.C., Lane W.S., Haber D.A., et al. BACH1, a novel helicase-like protein, interacts directly with BRCA1 and contributes to its DNA repair function. Cell. 2001;105:149–160. - PubMed
-
- Bridge W.L., Vandenberg C.J., Franklin R.J., Hiom K. The BRIP1 helicase functions independently of BRCA1 in the Fanconi anemia pathway for DNA crosslink repair. Nature Genet. 2005;37:953–957. - PubMed
-
- Litman R., Peng M., Jin Z., Zhang F., Zhang J., Powell S., Andreassen P.R., Cantor S.B. BACH1 is critical for homologous recombination and appears to be the Fanconi anemia gene product FANCJ. Cancer Cell. 2005;8:255–265. - PubMed
-
- Peng M., Litman R., Jin Z., Fong G., Cantor S.B. BACH1 is a DNA repair protein supporting BRCA1 damage response. Oncogene. 2006;25:2245–2253. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
