

Review
doi: 10.1098/rsbl.2006.0440.
Bare skin, blood and the evolution of primate colour vision
Affiliations
- PMID: 17148366
- PMCID: PMC1618887
- DOI: 10.1098/rsbl.2006.0440
Item in Clipboard
Review
Bare skin, blood and the evolution of primate colour vision
Biol Lett.
.
Abstract
We investigate the hypothesis that colour vision in primates was selected for discriminating the spectral modulations on the skin of conspecifics, presumably for the purpose of discriminating emotional states, socio-sexual signals and threat displays. Here we show that, consistent with this hypothesis, there are two dimensions of skin spectral modulations, and trichromats but not dichromats are sensitive to each. Furthermore, the M and L cone maximum sensitivities for routine trichromats are optimized for discriminating variations in blood oxygen saturation, one of the two blood-related dimensions determining skin reflectance. We also show that, consistent with the hypothesis, trichromat primates tend to be bare faced.
Figures
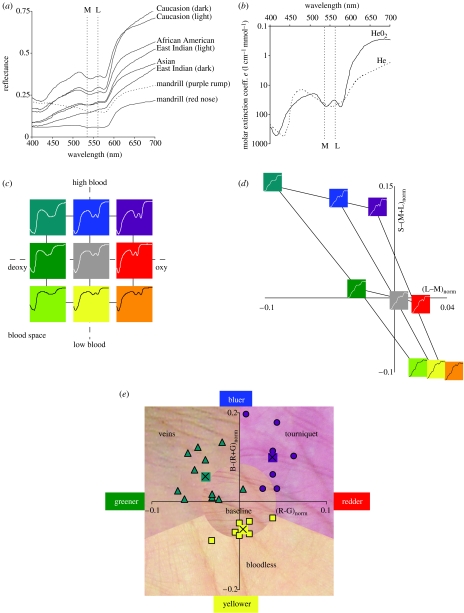
(a) Reflectance spectra from a variety of human skin (data from NCSU spectral database), and from one male primate, namely Mandrillus sphinx (Sumner & Mollon 2003). Also shown here and in (b) are the maximal sensitivities for the M and L cones for routine trichromats. (b) Absorption spectrum for oxygenated and deoxygenated haemoglobin (from Scott Prahl, Oregon Medical Laser Center, http://omlc.ogi.edu ). (c) Blood space for skin spectral modulation, showing the two principle variables that affect skin colour in the short term: haemoglobin oxygen saturation (x-axis), and haemoglobin skin concentration (y-axis). ‘High’, ‘baseline’ and ‘low’ values for these two variables were chosen, and the figure shows the nine skin spectra for all pairs of these parameter settings. Colours code the approximate direction of colour shift from baseline (centre). (d) Relative change from baseline for L−M and S−(L+M) for the nine model skin spectra varying over blood space from (c). Shown now are the filtered skin spectra actually reaching the retina. (e) Example skin colour modulations from modulations of blood variables. Data points show positions in this colour space for RGB values of skin under these conditions, along with the average values. See electronic supplementary material for the extended legend for this figure.
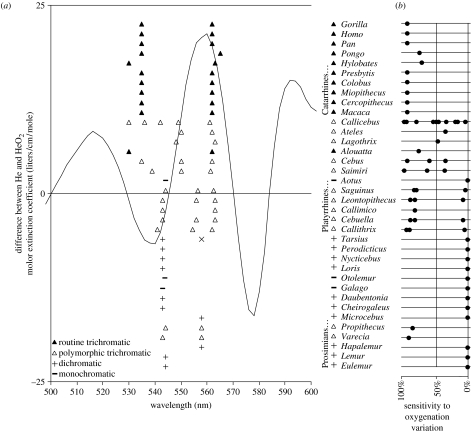
(a) The curve shows the difference between the absorption spectrum for oxygenated and deoxygenated haemoglobin. The difference between oxygenated and deoxygenated skin spectra lead to qualitatively identical curves, no matter the specific skin constants (i.e. same peak and valley wavelengths). Shown also are peak M/L cone sensitivities for the primate genera shown (reviewed in Surridge et al. (2003)). (Height in the plot for these is only to separate the data.) The vertical lines are at wavelengths where we would expect the maximal sensitivities of M and L cones to be, respectively, if they are optimally sensitive for oxygenation variation, and subject to the constraint that M and L jointly function as the single dichromat M/L cone. For Tarsius, Surridge et al. (2003) give two different values for the single M/L cone, and an ‘x’ point is shown for the longer wavelength one. For Cebuella, an allele at 543 nm has been added because Surridge & Mundy (2002, p. 2164) believe it probably exists but was not measured due to low sample size. (b) A plot of sensitivity of M/L cones to oxygenation variation, for each primate genus, where 100% would occur if the maximal sensitivities of M and L occurred at the optimum for oxygenation sensitivity. For polymorphic trichromatic primates, points are placed for each of the possible pairs of M/L cones. In several cases, Surridge et al. (2003) do not mention all three M/L cones, and we utilized the value from other genera in the same family. The line shows the average sensitivity for all M and L pairs centred around 543 nm (the typical dichromat maximal sensitivity wavelength), where M ranges as low as 500 nm.
References
-
- Allen G. Trubner & Co.; London, UK: 1879. The colour-sense: its origin and development.
-
- Hingston R.W.G. Edward Arnold; London, UK: 1933. The meaning of animal colour and adornment.
-
- Jablonski N.G, Chaplin G. The evolution of human skin coloration. J. Hum. Evol. 2000;39:57–106. doi:10.1006/jhev.2000.0403 - DOI - PubMed
-
- Jacobs G.H, Deegan J.F. Uniformity of colour vision in Old World monkeys. Proc. R. Soc. B. 1999;266:2023–2028. doi:10.1098/rspb.1999.0881 - DOI - PMC - PubMed
-
- Kienle A, Lilge L, Vitkin A, Patterson M.S, Wilson B.C, Hibst R, Steiner R. Why do veins appear blue? A new look at an old question. Appl. Opt. 1996;35:1151–1160. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases