

Development of the hemangioblast defines the onset of hematopoiesis in human ES cell differentiation cultures
- PMID: 17148580
- PMCID: PMC1852226
- DOI: 10.1182/blood-2006-09-047704
Development of the hemangioblast defines the onset of hematopoiesis in human ES cell differentiation cultures
Abstract
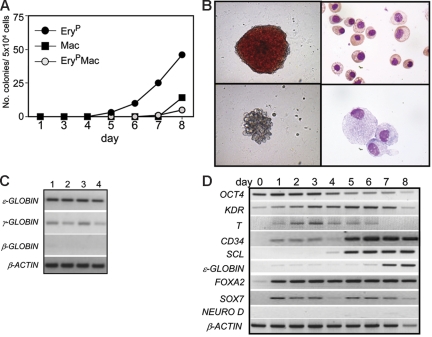
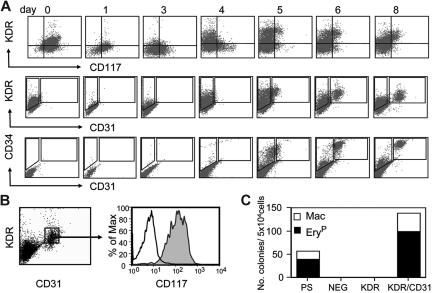
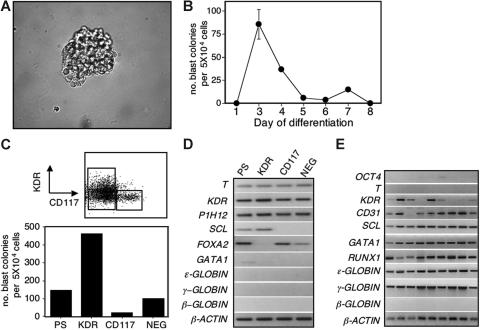
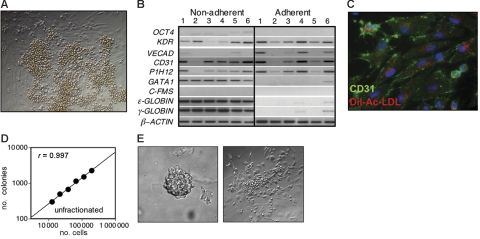
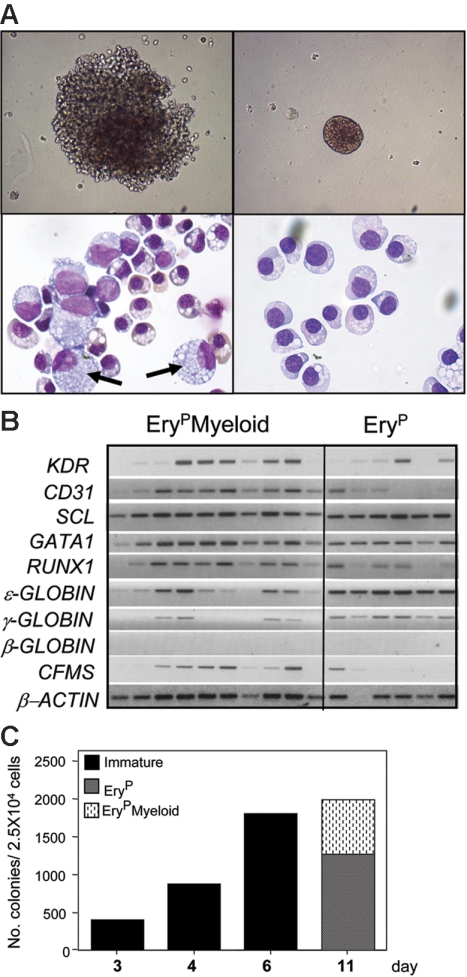
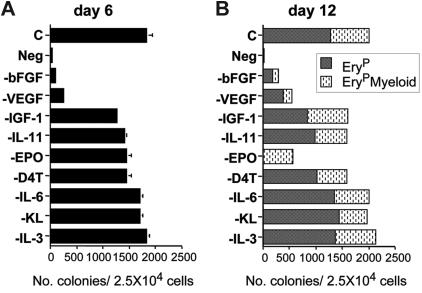
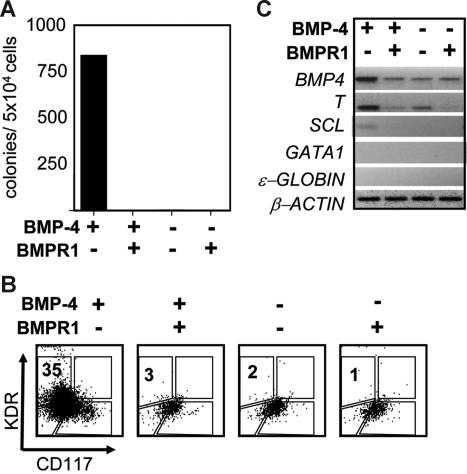
The onset of hematopoiesis in the mouse embryo and in the embryonic stem (ES) cell differentiation model is defined by the emergence of the hemangioblast, a progenitor with both hematopoietic and vascular potential. While there is evidence for the existence of a hemangioblast in the mouse, it is unclear if this progenitor develops during the establishment of the human hematopoietic system. In this report, we have mapped hematopoietic development in human ES cell (hESC) differentiation cultures and demonstrated that a comparable hemangioblast population exists. The human hemangioblasts were identified by their capacity to generate blast colonies that display both hematopoietic and vascular potential. These colony-forming cells express the receptor tyrosine kinase KDR (VEGF receptor 2) and represent a transient population that develops in BMP-4-stimulated embryoid bodies (EBs) between 72 and 96 hours of differentiation, prior to the onset of the primitive erythroid program. Two distinct types of hemangioblasts were identified, those that give rise to primitive erythroid cells, macrophages, and endothelial cells and those that generate only the primitive erythroid population and endothelial cells. These findings demonstrate for the first time the existence of the human hemangioblast and in doing so identify the earliest stage of hematopoietic commitment.
Figures
References
-
- Haar JL, Ackerman GA. A phase and electron microscopic study of vasculogenesis and erythropoiesis in the yolk sac of the mouse. Anat Rec. 1971;170:199–224. - PubMed
-
- Palis J, McGrath KE, Kingsley PD. Initiation of hematopoiesis and vasculogenesis in murine yolk sac explants. Blood. 1995;86:156–163. - PubMed
-
- Palis J, Roberston S, Kennedy M, Wall C, Keller G. Development of erythroid and myeloid progenitors in the yolk sac and embryo proper of the mouse. Development. 1999;126:5073–5084. - PubMed
-
- Sabin FR. Studies on the origin of blood vessels and of red corpuscles as seen in the living blastoderm of the chick during the second day of incubation. Contrib Embryol. 1920;9:213–262.
-
- Murray PDF. The development in vitro of the blood of the early chick embryo. Proc Royal Soc London. 1932;11:497–521.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
