

MAFA controls genes implicated in insulin biosynthesis and secretion
- PMID: 17149590
- PMCID: PMC2196442
- DOI: 10.1007/s00125-006-0490-2
MAFA controls genes implicated in insulin biosynthesis and secretion
Abstract
Aims/hypothesis: Effects of the transcription factor v-maf musculoaponeurotic fibrosarcoma oncogene homologue A (MAFA) on the regulation of beta cell gene expression and function were investigated.
Materials and methods: INS-1 stable cell lines permitting inducible up- or downregulation of this transcription factor were established.
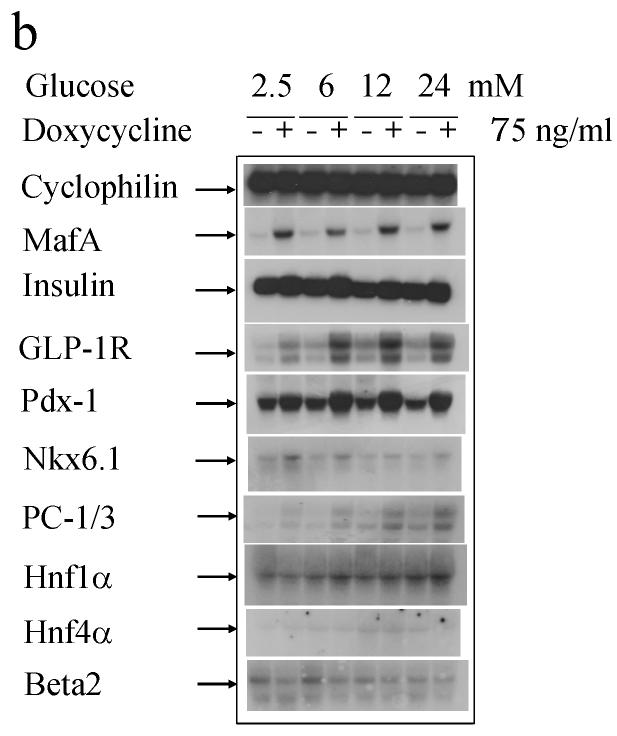
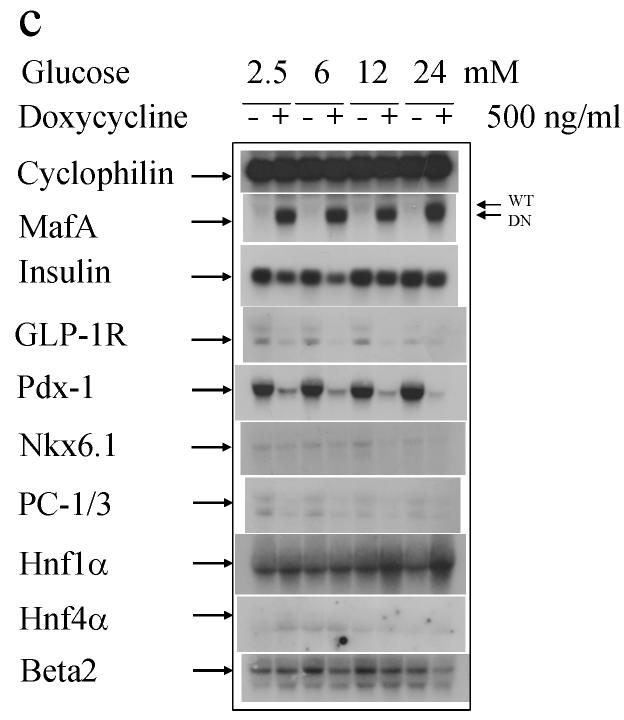
Results: MAFA overproduction enhanced and its dominant-negative mutant (DN-MAFA) diminished binding of the factor to the insulin promoter, correlating with insulin mRNA levels and cellular protein content. Glucose-stimulated insulin secretion was facilitated by MAFA and blunted by DN-MAFA. This is partly due to alterations in glucokinase production, the glucose sensor of beta cells. In addition, the expression of important beta cell genes, e.g. those encoding solute carrier family 2 (facilitated glucose transporter), member 2 (formerly known as GLUT2), pancreatic and duodenal homeobox factor 1 (PDX1), NK6 transcription factor-related, locus 1 (NKX6-1), glucagon-like peptide 1 receptor (GLP1R), prohormone convertase 1/3 (PCSK1) and pyruvate carboxylase (PC), was regulated positively by MAFA and negatively by DN-MAFA.
Conclusions/interpretation: The data suggest that MAFA is not only a key activator of insulin transcription, but also a master regulator of genes implicated in maintaining beta cell function, in particular metabolism-secretion coupling, proinsulin processing and GLP1R signalling. Our in vitro study provides molecular targets that explain the phenotype of recently reported Mafa-null mice. We also demonstrate that MAFA is produced specifically in beta cells of human islets. Glucose influenced DNA-binding activity of MAFA in rat islets in a bell-shaped manner. MAFA thus qualifies as a master regulator of beta-cell-specific gene expression and function.
Figures













References
-
- Naya FJ, Stellrecht CM, Tsai MJ. Tissue-specific regulation of the insulin gene by a novel basic helix-loop-helix transcription factor. Genes Dev. 1995;9:1009–1019. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
