

Effects of introgression and recombination on haplotype structure and linkage disequilibrium surrounding a locus encoding Bymovirus resistance in barley
- PMID: 17151251
- PMCID: PMC1800611
- DOI: 10.1534/genetics.106.063800
Effects of introgression and recombination on haplotype structure and linkage disequilibrium surrounding a locus encoding Bymovirus resistance in barley
Abstract
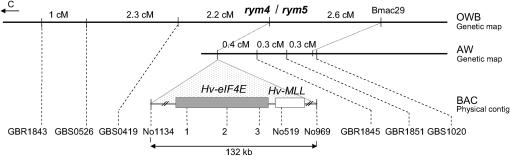
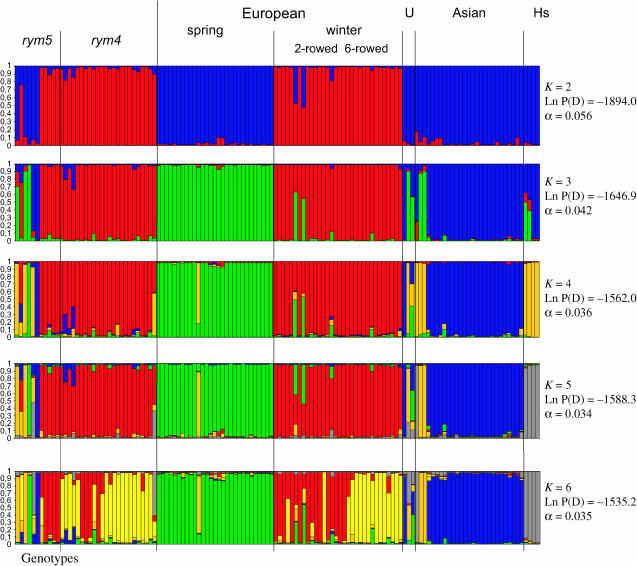
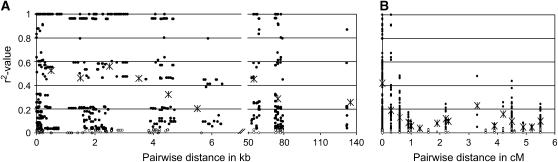
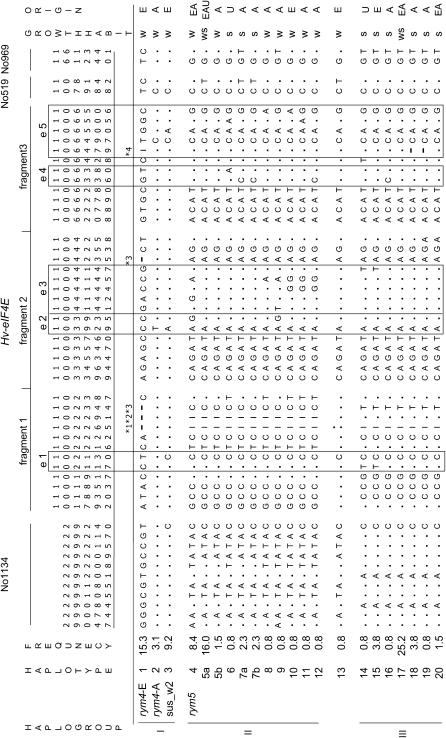
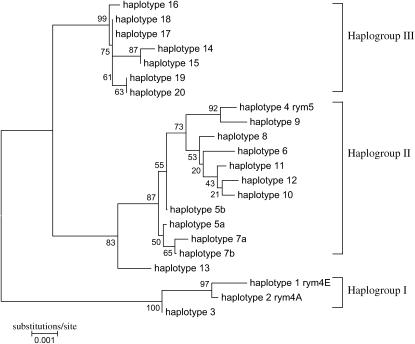
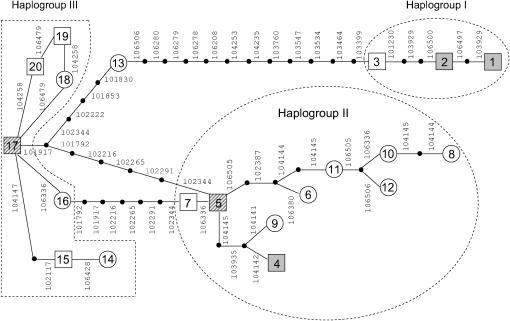
We present a detailed analysis of linkage disequilibrium (LD) in the physical and genetic context of the barley gene Hv-eIF4E, which confers resistance to the barley yellow mosaic virus (BYMV) complex. Eighty-three SNPs distributed over 132 kb of Hv-eIF4E and six additional fragments genetically mapped to its flanking region were used to derive haplotypes from 131 accessions. Three haplogroups were recognized, discriminating between the alleles rym4 and rym5, which each encode for a spectrum of resistance to BYMV. With increasing map distance, haplotypes of susceptible genotypes displayed diverse patterns driven mainly by recombination, whereas haplotype diversity within the subgroups of resistant genotypes was limited. We conclude that the breakdown of LD within 1 cM of the resistance gene was generated mainly by susceptible genotypes. Despite the LD decay, a significant association between haplotype and resistance to BYMV was detected up to a distance of 5.5 cM from the resistance gene. The LD pattern and the haplotype structure of the target chromosomal region are the result of interplay between low recombination and recent breeding history.
Figures

References
-
- Abdel-Ghani, A. H., H. K. Parzies, S. Ceccarelli, S. Grando and H. H. Geiger, 2005. Estimation of quantitative genetic parameters for outcrossing-related traits in barley. Crop Sci. 45: 98–105.
-
- Akey, J. M., K. Zhang, M. M. Xiong and L. Jin, 2003. The effect of single nucleotide polymorphism identification strategies on estimates of linkage disequilibrium. Mol. Biol. Evol. 20: 232–242. - PubMed
-
- Anderson, J. A., G. A. Churchill, J. E. Autrique, S. D. Tanksley and M. E. Sorrells, 1993. Optimizing parental selection for genetic-linkage maps. Genome 36: 181–186. - PubMed
-
- Badr, A., K. Muller, R. Schafer-Pregl, H. El Rabey, S. Effgen et al., 2000. On the origin and domestication history of barley (Hordeum vulgare). Mol. Biol. Evol. 17: 499–510. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
