

Novel retrotransposon analysis reveals multiple mobility pathways dictated by hosts
- PMID: 17151346
- PMCID: PMC1716264
- DOI: 10.1101/gr.5542607
Novel retrotransposon analysis reveals multiple mobility pathways dictated by hosts
Abstract
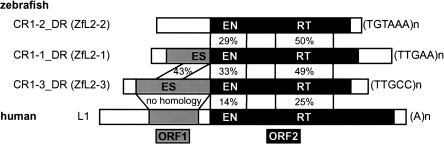
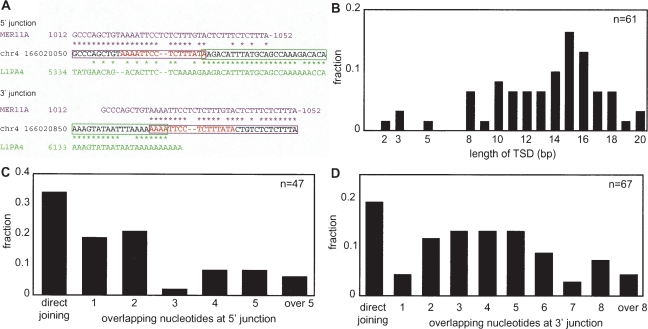
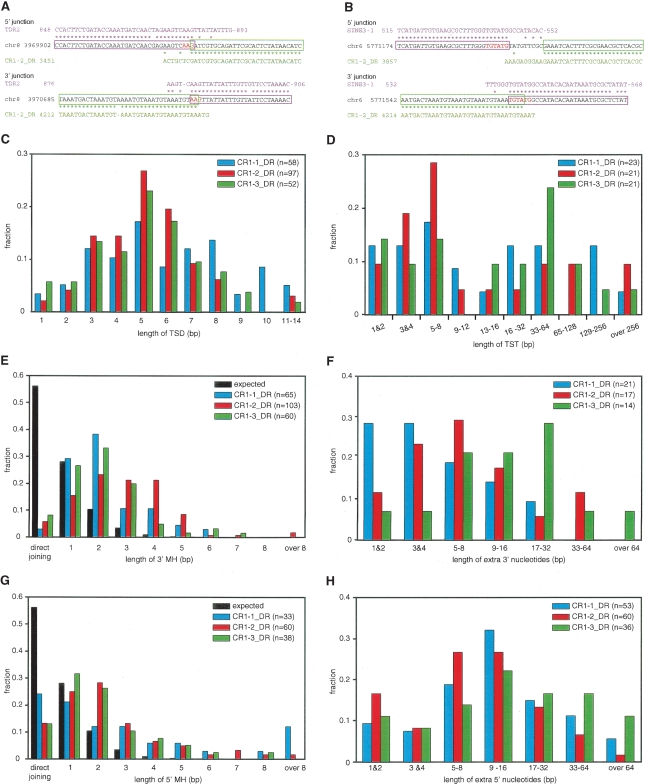
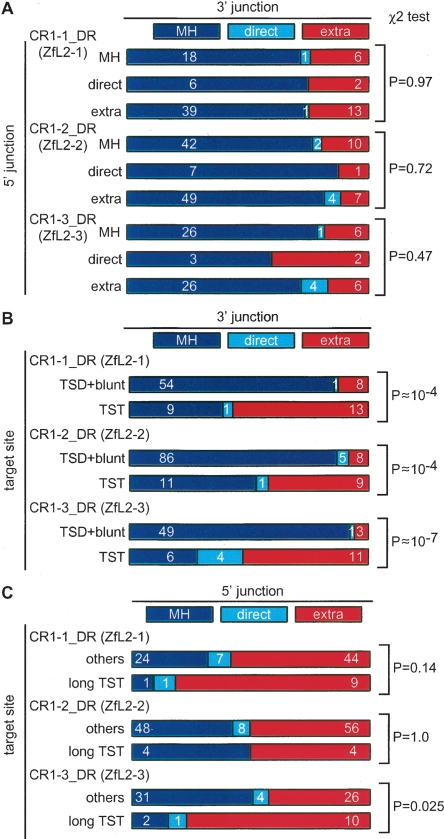
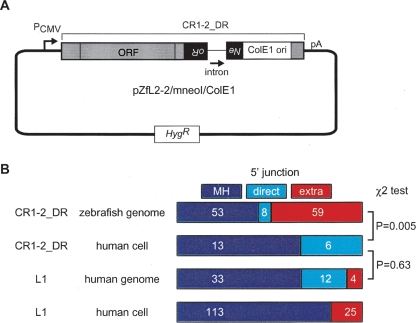
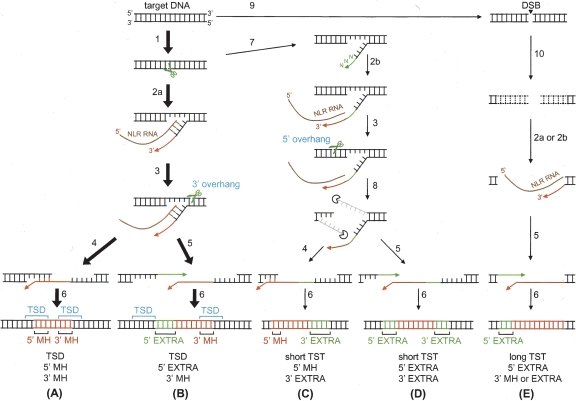
Autonomous non-long-terminal-repeat retrotransposons (NLRs) proliferate by retrotransposition via coordinated reactions of target DNA cleavage and reverse transcription by a mechanism called target-primed reverse transcription (TPRT). Whereas this mechanism guarantees the covalent attachment of the NLR and its target site at the 3' junction, mechanisms for the joining at the 5' junction have been conjectural. To better understand the retrotransposition pathways, we analyzed target-NLR junctions of zebrafish NLRs with a new method of identifying genomic copies that reside within other transposons, termed "target analysis of nested transposons" (TANT). Application of the TANT method revealed various features of the zebrafish NLR integrants; for example, half of the integrants carry extra nucleotides at the 5' junction, which is in stark contrast to the major human NLR, LINE-1. Interestingly, in a cell culture assay, retrotransposition of the zebrafish NLR in heterologous human cells did not bear extra 5' nucleotides, indicating that the choice of the 5' joining pathway is affected by the host. Our results suggest that several pathways exist for NLR retrotransposition and argue in favor of host protein involvement. With genomic sequence information accumulating exponentially, our data demonstrate the general applicability of the TANT method for the analysis of a wide variety of retrotransposons.
Figures
Similar articles
-
Mobility pathways for vertebrate L1, L2, CR1, and RTE clade retrotransposons.Mol Biol Evol. 2008 Jun;25(6):1148-57. doi: 10.1093/molbev/msn061. Epub 2008 Mar 14. Mol Biol Evol. 2008. PMID: 18343891
-
Genomic alterations upon integration of zebrafish L1 elements revealed by the TANT method.Gene. 2006 Nov 15;383:108-16. doi: 10.1016/j.gene.2006.07.038. Epub 2006 Aug 23. Gene. 2006. PMID: 17049188
-
A new mechanism to ensure integration during LINE retrotransposition: a suggestion from analyses of the 5' extra nucleotides.Gene. 2012 Sep 1;505(2):345-51. doi: 10.1016/j.gene.2012.02.047. Epub 2012 Mar 1. Gene. 2012. PMID: 22405943
-
DIRS-1 and the other tyrosine recombinase retrotransposons.Cytogenet Genome Res. 2005;110(1-4):575-88. doi: 10.1159/000084991. Cytogenet Genome Res. 2005. PMID: 16093711 Review.
-
Crossing the LINE Toward Genomic Instability: LINE-1 Retrotransposition in Cancer.Front Chem. 2015 Dec 16;3:68. doi: 10.3389/fchem.2015.00068. eCollection 2015. Front Chem. 2015. PMID: 26734601 Free PMC article. Review.
Cited by
-
Endonuclease-independent insertion provides an alternative pathway for L1 retrotransposition in the human genome.Nucleic Acids Res. 2007;35(11):3741-51. doi: 10.1093/nar/gkm317. Epub 2007 May 21. Nucleic Acids Res. 2007. PMID: 17517773 Free PMC article.
-
The fifth Japanese meeting on biological function and evolution through interactions between hosts and transposable elements.Mob DNA. 2022 Jan 13;13(1):3. doi: 10.1186/s13100-022-00261-7. Mob DNA. 2022. PMID: 35027075 Free PMC article.
-
Cross-Kingdom Commonality of a Novel Insertion Signature of RTE-Related Short Retroposons.Genome Biol Evol. 2018 Jun 1;10(6):1471-1483. doi: 10.1093/gbe/evy098. Genome Biol Evol. 2018. PMID: 29850801 Free PMC article.
-
A mobile threat to genome stability: The impact of non-LTR retrotransposons upon the human genome.Semin Cancer Biol. 2010 Aug;20(4):211-21. doi: 10.1016/j.semcancer.2010.03.001. Epub 2010 Mar 20. Semin Cancer Biol. 2010. PMID: 20307669 Free PMC article. Review.
-
Genetic evidence that the non-homologous end-joining repair pathway is involved in LINE retrotransposition.PLoS Genet. 2009 Apr;5(4):e1000461. doi: 10.1371/journal.pgen.1000461. Epub 2009 Apr 24. PLoS Genet. 2009. PMID: 19390601 Free PMC article.
References
-
- Babushok D.V., Ostertag E.M., Courtney C.E., Choi J.M., Kazazian H.H., Jr., Ostertag E.M., Courtney C.E., Choi J.M., Kazazian H.H., Jr., Courtney C.E., Choi J.M., Kazazian H.H., Jr., Choi J.M., Kazazian H.H., Jr., Kazazian H.H., Jr. L1 integration in a transgenic mouse model. Genome Res. 2006;16:240–250. - PMC - PubMed
-
- Coros C.J., Landthaler M., Piazza C.L., Beauregard A., Esposito D., Perutka J., Lambowitz A.M., Belfort M., Landthaler M., Piazza C.L., Beauregard A., Esposito D., Perutka J., Lambowitz A.M., Belfort M., Piazza C.L., Beauregard A., Esposito D., Perutka J., Lambowitz A.M., Belfort M., Beauregard A., Esposito D., Perutka J., Lambowitz A.M., Belfort M., Esposito D., Perutka J., Lambowitz A.M., Belfort M., Perutka J., Lambowitz A.M., Belfort M., Lambowitz A.M., Belfort M., Belfort M. Retrotransposition strategies of the Lactococcus lactis Ll.LtrB group II intron are dictated by host identity and cellular environment. Mol. Microbiol. 2005;56:509–524. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources