

Valproic acid and butyrate induce apoptosis in human cancer cells through inhibition of gene expression of Akt/protein kinase B
- PMID: 17156483
- PMCID: PMC1762018
- DOI: 10.1186/1476-4598-5-71
Valproic acid and butyrate induce apoptosis in human cancer cells through inhibition of gene expression of Akt/protein kinase B
Abstract
Background: In eukaryotic cells, the genomic DNA is packed with histones to form the nucleosome and chromatin structure. Reversible acetylation of the histone tails plays an important role in the control of specific gene expression. Mounting evidence has established that histone deacetylase inhibitors selectively induce cellular differentiation, growth arrest and apoptosis in variety of cancer cells, making them a promising class of anticancer drugs. However, the molecular mechanisms of the anti-cancer effects of these inhibitors have yet to be understood.
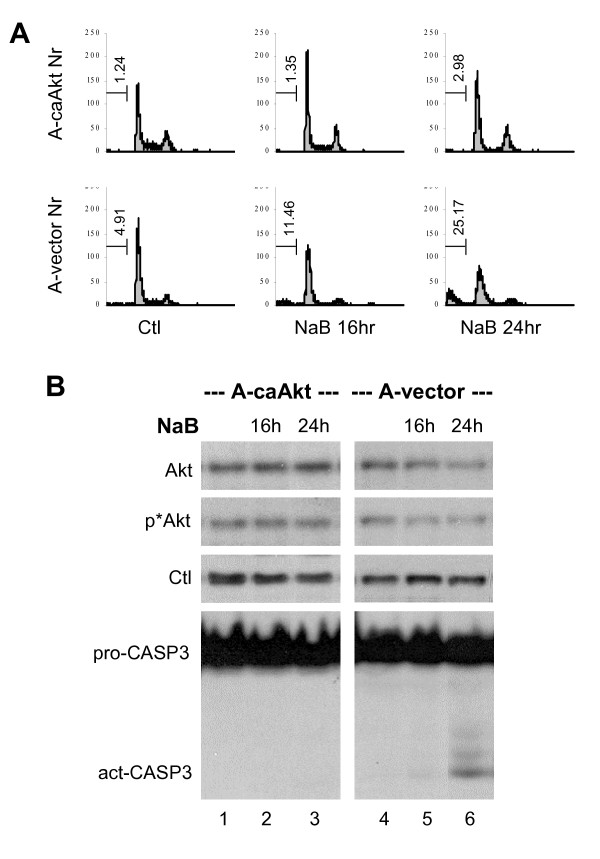
Results: Here, we report that a key determinant for the susceptibility of cancer cells to histone deacetylase inhibitors is their ability to maintain cellular Akt activity in response to the treatment. Also known as protein kinase B, Akt is an essential pro-survival factor in cell proliferation and is often deregulated during tumorigenesis. We show that histone deacetylase inhibitors, such as valproic acid and butyrate, impede Akt1 and Akt2 expression, which leads to Akt deactivation and apoptotic cell death. In addition, valproic acid and butyrate induce apoptosis through the caspase-dependent pathway. The activity of caspase-9 is robustly activated upon valproic acid or butyrate treatment. Constitutively active Akt is able to block the caspase activation and rescues cells from butyrate-induced apoptotic cell death.
Conclusion: Our study demonstrates that although the primary target of histone deacetylase inhibitors is transcription, it is the capacity of cells to maintain cellular survival networks that determines their fate of survival.
Figures





References
-
- Wolffe AP. Histones, nucleosomes and the roles of chromatin structure in transcriptional control. Biochem Soc Trans. 1997;25:354–358. - PubMed
-
- Li Q, Wrange O. Translational positioning of a nucleosomal glucocorticoid response element modulates glucocorticoid receptor affinity. Genes Dev. 1993;7:2471–2482. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
