

The Saccharomyces cerevisiae YFR041C/ERJ5 gene encoding a type I membrane protein with a J domain is required to preserve the folding capacity of the endoplasmic reticulum
- PMID: 17157937
- PMCID: PMC1847348
- DOI: 10.1016/j.bbamcr.2006.10.011
The Saccharomyces cerevisiae YFR041C/ERJ5 gene encoding a type I membrane protein with a J domain is required to preserve the folding capacity of the endoplasmic reticulum
Abstract
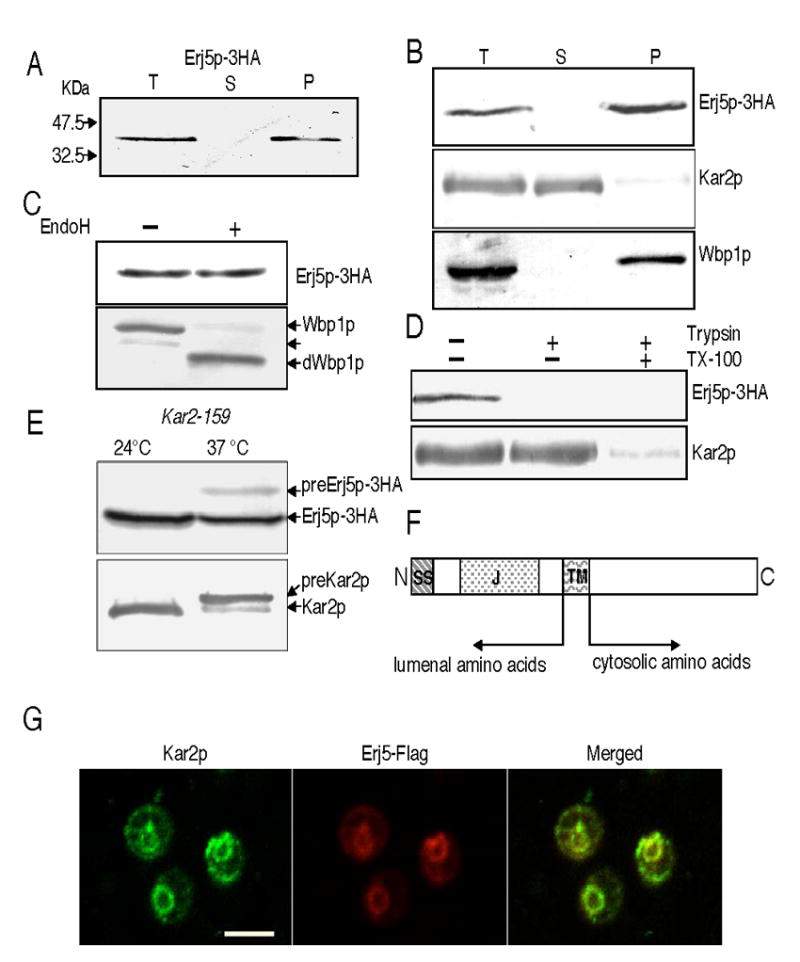
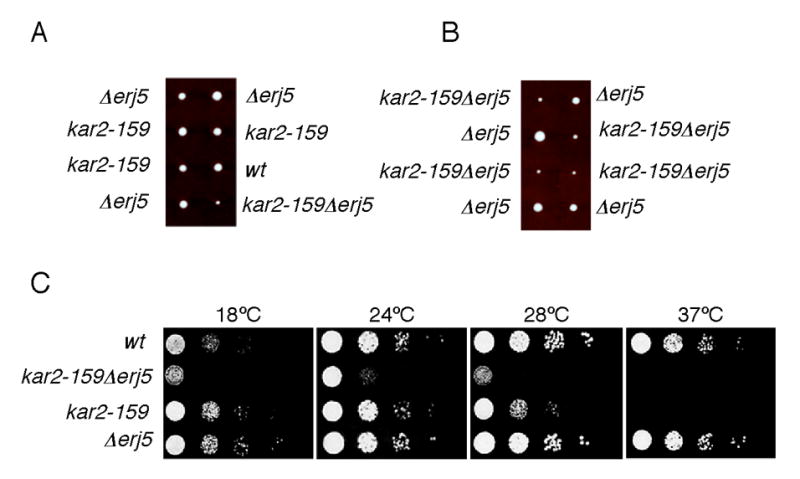
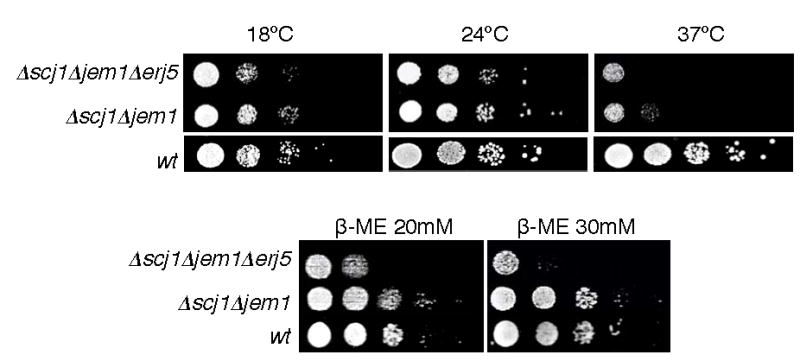
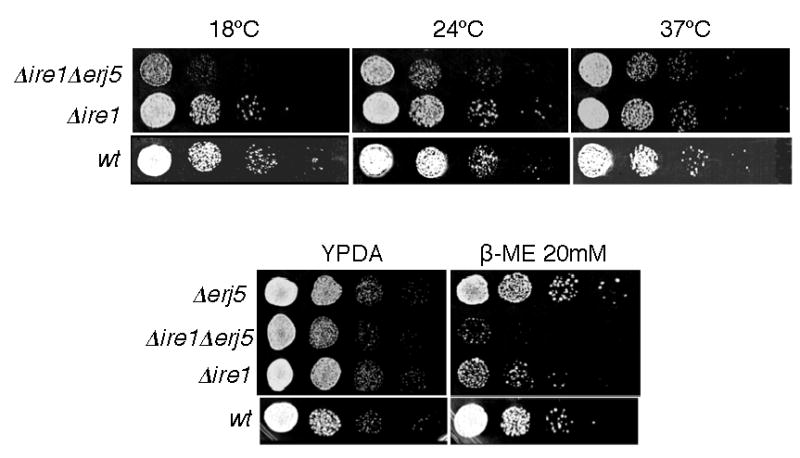
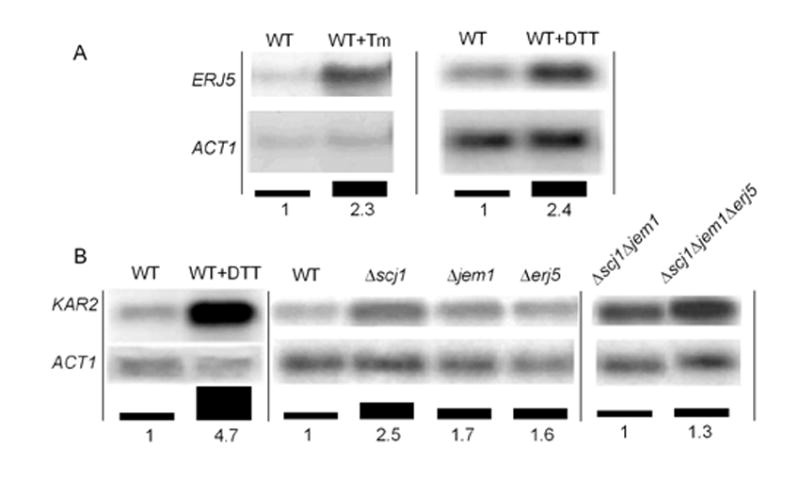
YFR041C/ERJ5 was identified in Saccharomyces cerevisiae as a gene regulated by the unfolded protein response pathway (UPR). The open reading frame of the gene has a J domain characteristic of the DnaJ chaperone family of proteins that regulate the activity of Hsp70 chaperones. We determined the expression and topology of Erj5p, a type I membrane protein with a J domain in the lumen of the endoplasmic reticulum (ER) that colocalizes with Kar2p, the major Hsp70 in the yeast ER. We identified synthetic interactions of Deltaerj5 with mutations in genes involved in protein folding in the ER (kar2-159, Deltascj1Deltajem1) and in the induction of the unfolded protein response (Deltaire1). Loss of Erj5p in yeast cells with impaired ER protein folding capacity increased sensitivity to agents that cause ER stress. We identified the ERJ5 mRNA and confirmed that agents that promote accumulation of misfolded proteins in the ER regulate its abundance. We found that loss of the non-essential ERJ5 gene leads to a constitutively induced UPR, indicating that ERJ5 is required for maintenance of an optimal folding environment in the yeast ER.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
