

The anaphase-promoting complex/cyclosome is required for anaphase progression in multinucleated Ashbya gossypii cells
- PMID: 17158735
- PMCID: PMC1797942
- DOI: 10.1128/EC.00364-06
The anaphase-promoting complex/cyclosome is required for anaphase progression in multinucleated Ashbya gossypii cells
Abstract
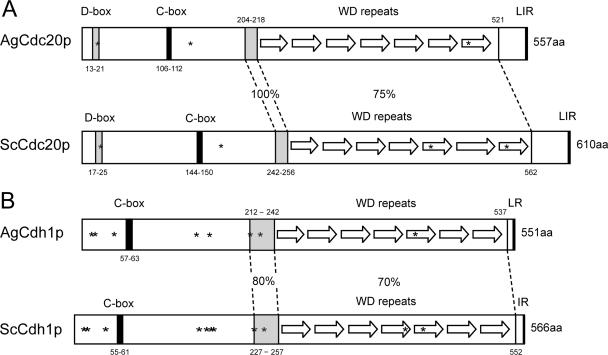
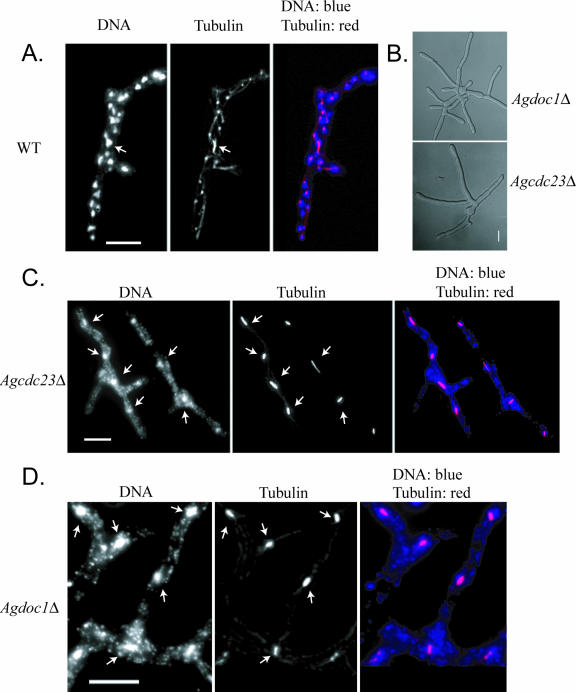
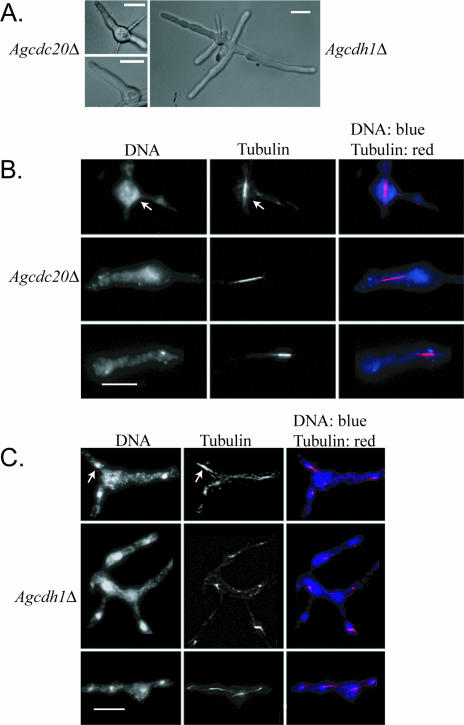
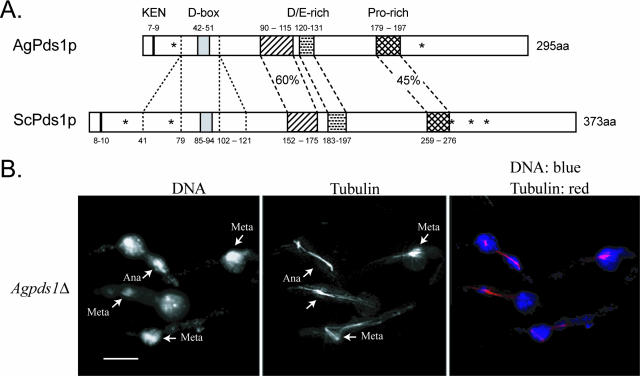
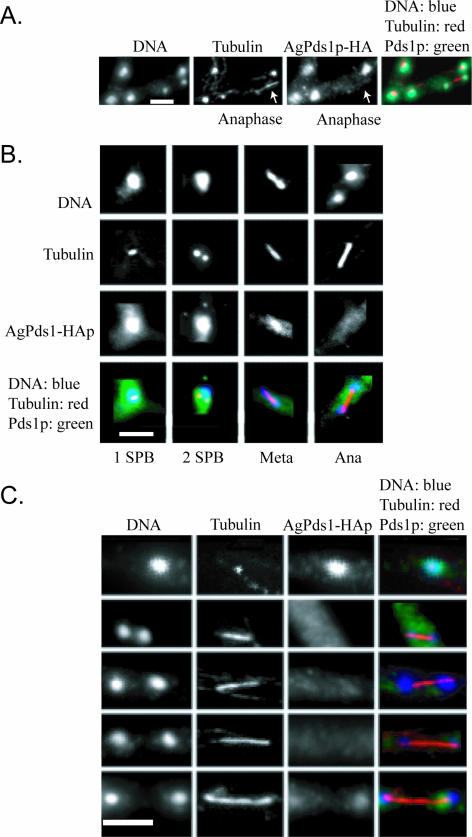
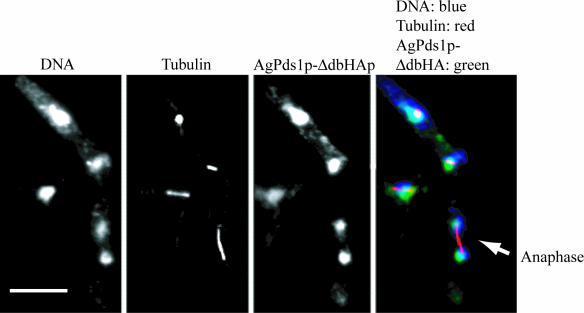
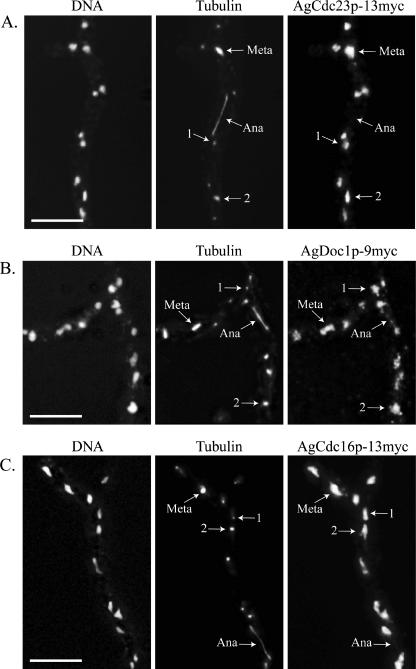
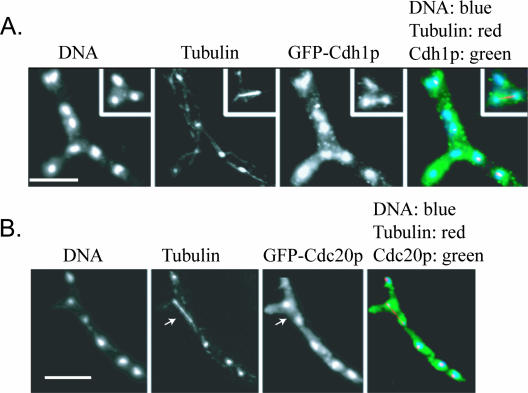
Regulated protein degradation is essential for eukaryotic cell cycle progression. The anaphase-promoting complex/cyclosome (APC/C) is responsible for the protein destruction required for the initiation of anaphase and the exit from mitosis, including the degradation of securin and B-type cyclins. We initiated a study of the APC/C in the multinucleated, filamentous ascomycete Ashbya gossypii to understand the mechanisms underlying the asynchronous mitosis observed in these cells. These experiments were motivated by previous work which demonstrated that the mitotic cyclin AgClb1/2p persists through anaphase, suggesting that the APC/C may not be required for the division cycle in A. gossypii. We have now found that the predicted APC/C components AgCdc23p and AgDoc1p and the targeting factors AgCdc20p and AgCdh1p are essential for growth and nuclear division. Mutants lacking any of these factors arrest as germlings with nuclei blocked in mitosis. A likely substrate of the APC/C is the securin homologue AgPds1p, which is present in all nuclei in hyphae except those in anaphase. The destruction box sequence of AgPds1p is required for this timed disappearance. To investigate how the APC/C may function to degrade AgPds1p in only the subset of anaphase nuclei, we localized components and targeting subunits of the APC/C. Remarkably, AgCdc23p, AgDoc1p, and AgCdc16p were found in all nuclei in all cell cycle stages, as were the APC/C targeting factors AgCdc20p and AgCdh1p. These data suggest that the AgAPC/C may be constitutively active across the cell cycle and that proteolysis in these multinucleated cells may be regulated at the level of substrates rather than by the APC/C itself.
Figures
References
-
- Alberti-Segui, C., F. Dietrich, R. Altmann-Johl, D. Hoepfner, and P. Philippsen. 2001. Cytoplasmic dynein is required to oppose the force that moves nuclei towards the hyphal tip in the filamentous ascomycete Ashbya gossypii. J. Cell Sci. 114:975-986. - PubMed
-
- Altmann-Jöhl, R., and P. Philippsen. 1996. AgTHR4, a new selection marker for transformation of the filamentous fungus Ashbya gossypii, maps in a four-gene cluster that is conserved between A. gossypii and Saccharomyces cerevisiae. Mol. Gen. Genet. 250:69-80. - PubMed
-
- Ayad-Durieux, Y., P. Knechtle, S. Goff, F. Dietrich, and P. Philippsen. 2000. A PAK-like protein kinase is required for maturation of young hyphae and septation in the filamentous ascomycete Ashbya gossypii. J. Cell Sci. 113:4563-4575. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
