

MDC1 accelerates nonhomologous end-joining of dysfunctional telomeres
- PMID: 17158742
- PMCID: PMC1686600
- DOI: 10.1101/gad.1496606
MDC1 accelerates nonhomologous end-joining of dysfunctional telomeres
Abstract
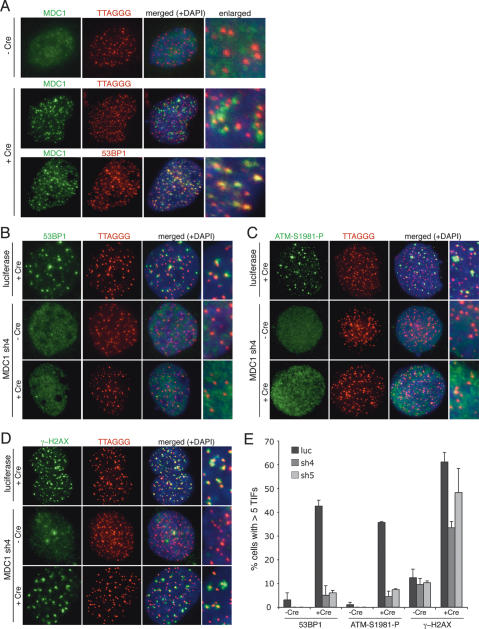
Here we document the role of MDC1 (mediator of DNA damage checkpoint 1) in the detection and repair of human and mouse telomeres rendered dysfunctional through inhibition of TRF2. Consistent with its role in promoting DNA damage foci, MDC1 knockdown affected the formation of telomere dysfunction-induced foci (TIFs), diminishing the accumulation of phosphorylated ATM, 53BP1, Nbs1, and to a lesser extent, gamma-H2AX. In addition to this effect on TIFs, the rate of nonhomologous end-joining (NHEJ) of dysfunctional telomeres was significantly decreased when MDC1 itself or its recruitment to chromatin was inhibited. MDC1 appeared to promote a step in the NHEJ pathway after the removal of the 3' telomeric overhang. The acceleration of NHEJ was unlikely to be due to increased presence of 53BP1 and Mre11 in TIFs, since knockdown of neither factor affected telomere fusions. Furthermore, relevant cell cycle effectors (Chk2, p53, and p21) of the ATM kinase pathway were unaffected and there was no change in the rate of cell cycle progression. We propose that the binding of MDC1 to gamma-H2AX directly affects NHEJ in a manner that is independent of the ATM-dependent cell cycle arrest pathway.
Figures



References
-
- Celli, G., de Lange, T. DNA processing not required for ATM-mediated telomere damage response after TRF2 deletion. Nat. Cell Biol. 2005;7:712–718. - PubMed
-
- Celli, G.B., Lazzerini Denchi, E., de Lange, T. Ku70 stimulates fusion of dysfunctional telomeres yet protects chromosome ends from homologous recombination. Nat. Cell Biol. 2006;8:885–890. - PubMed
-
- di d’Adda Fagagna, F., Reaper, P.M., Clay-Farrace, L., Fiegler, H., Carr, P., Von Zglinicki, T., Saretzki, G., Carter, N.P., Jackson, S.P. A DNA damage checkpoint response in telomere-initiated senescence. Nature. 2003;426:194–198. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous