

Pin1 stabilizes Emi1 during G2 phase by preventing its association with SCF(betatrcp)
- PMID: 17159919
- PMCID: PMC1796751
- DOI: 10.1038/sj.embor.7400853
Pin1 stabilizes Emi1 during G2 phase by preventing its association with SCF(betatrcp)
Abstract
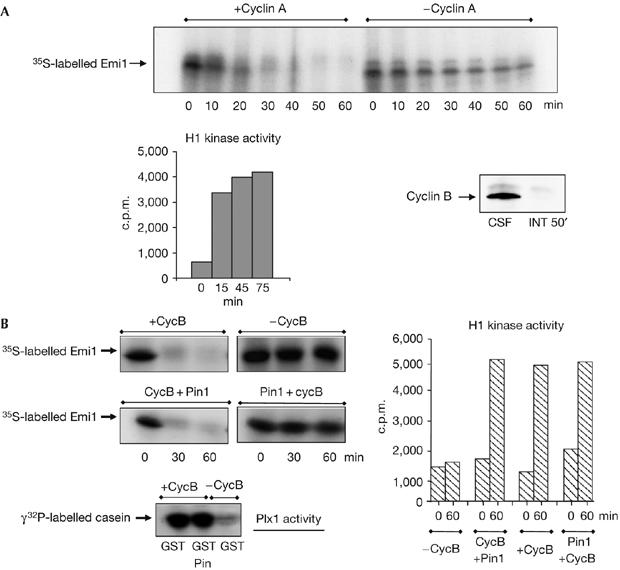
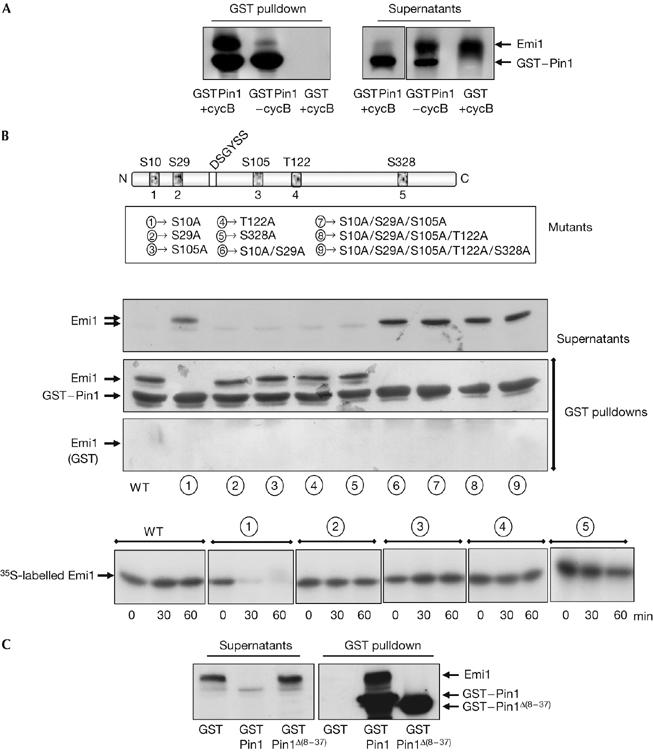
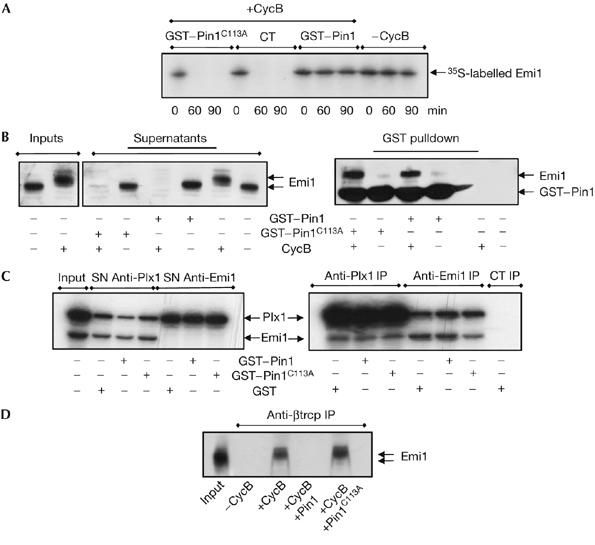
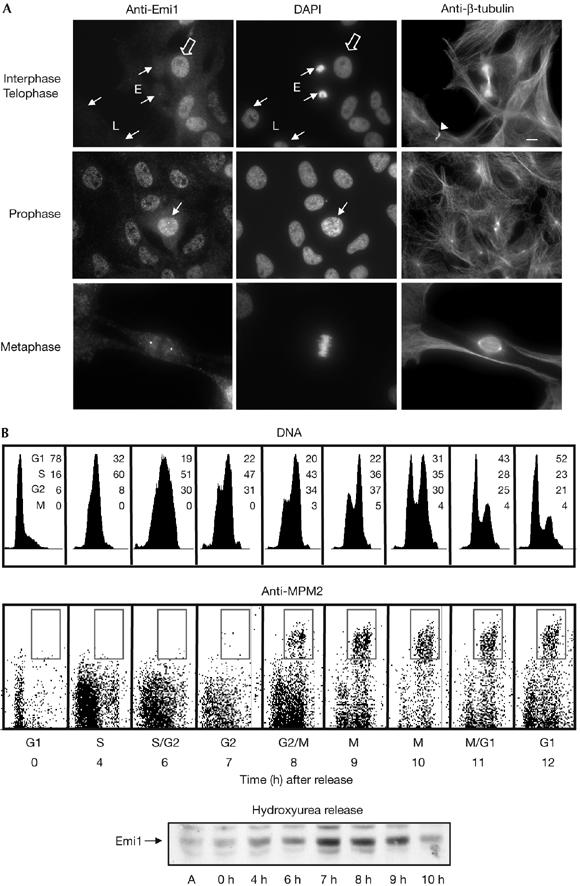
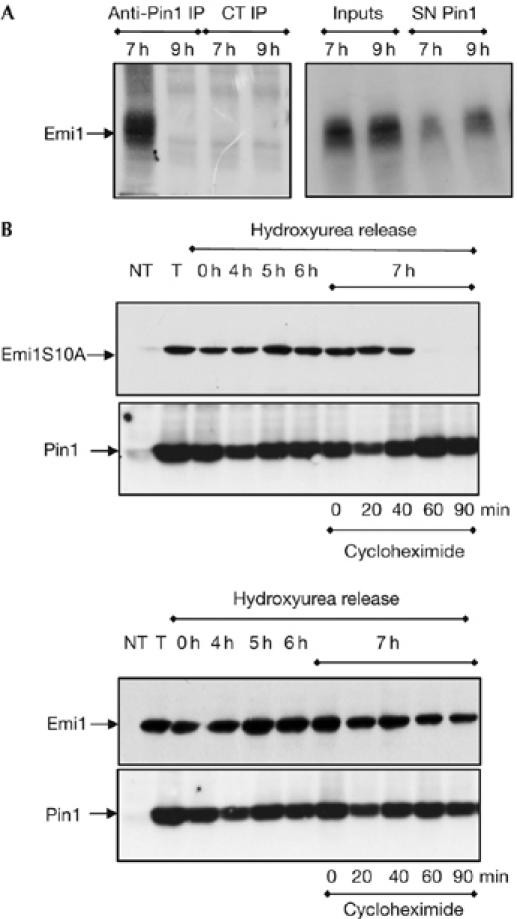
The anaphase-promoting complex (APC) early mitotic inhibitor 1 (Emi1) is required to induce S- and M-phase entries by stimulating the accumulation of cyclin A and cyclin B through APC(Cdh1/cdc20) inhibition. In this report, we show that Emi1 proteolysis can be induced by cyclin A/cdk (cdk for cyclin-dependent kinase). Paradoxically, Emi1 is stable during G2 phase, when cyclin A/cdk, Plx1 and SCF(betatrcp) (SCF for Skp1-Cul1-Fbox protein)--which play a role in its degradation--are active. Here, we identify Pin1 as a new regulator of Emi1 that induces Emi1 stabilization by preventing its association with SCF(betatrcp). We show that Pin1 binds to Emi1 and prevents its association with betatrcp in an isomerization-dependent pathway. We also show that Emi1-Pin1 binding is present in vivo in XL2 cells during G2 phase and that this association protects Emi1 from being degraded during this phase of the cell cycle. We propose that S- and M-phase entries are mediated by the accumulation of cyclin A and cyclin B through a Pin1-dependent stabilization of Emi1 during G2.
Figures
References
-
- Castro A, Bernis C, Vigneron S, Labbe JC, Lorca T (2005) The anaphase-promoting complex: a key factor in the regulation of cell cycle. Oncogene 24: 314–325 - PubMed
-
- Eldridge AG, Loktev AV, Hansen DV, Verschuren EW, Reimann JD, Jackson PK (2006) The Evi5 oncogene regulates cyclin accumulation by stabilizing the anaphase-promoting complex inhibitor Emi1. Cell 27: 367–380 - PubMed
-
- Guardavaccaro D, Kudo Y, Boulaire J, Barchi M, Busino L, Donzelli M, Margottin-Goguet F, Jackson PK, Yamasaki L, Pagano M (2003) Control of meiotic and mitotic progression by the F box protein β-Trcp1 in vivo. Dev Cell 4: 799–812 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
