

An asymmetric underlying rule in the assignment of codons: possible clue to a quick early evolution of the genetic code via successive binary choices
- PMID: 17164478
- PMCID: PMC1781368
- DOI: 10.1261/rna.257607
An asymmetric underlying rule in the assignment of codons: possible clue to a quick early evolution of the genetic code via successive binary choices
Abstract
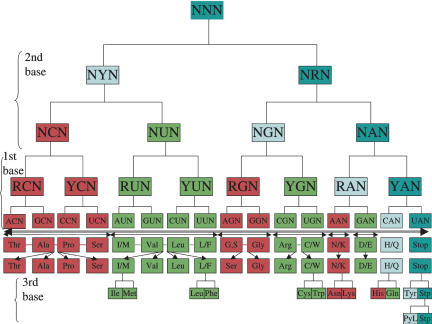
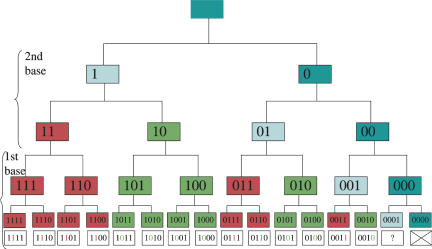
Aminoacyl-tRNA synthetases (aaRSs) are responsible for creating the pool of correctly charged aminoacyl-tRNAs that are necessary for the translation of genetic information (mRNA) by the ribosome. Each aaRS belongs to either one of only two classes with two different mechanisms of aminoacylation, making use of either the 2'OH (Class I) or the 3'OH (Class II) of the terminal A76 of the tRNA and approaching the tRNA either from the minor groove (2'OH) or the major groove (3'OH). Here, an asymmetric pattern typical of differentiation is uncovered in the partition of the codon repertoire, as defined by the mechanism of aminoacylation of each corresponding tRNA. This pattern can be reproduced in a unique cascade of successive binary decisions that progressively reduces codon ambiguity. The deduced order of differentiation is manifestly driven by the reduction of translation errors. A simple rule can be defined, decoding each codon sequence in its binary class, thereby providing both the code and the key to decode it. Assuming that the partition into two mechanisms of tRNA aminoacylation is a relic that dates back to the invention of the genetic code in the RNA World, a model for the assignment of amino acids in the codon table can be derived. The model implies that the stop codon was always there, as the codon whose tRNA cannot be charged with any amino acid, and makes the prediction of an ultimate differentiation step, which is found to correspond to the codon assignment of the 22nd amino acid pyrrolysine in archaebacteria.
Figures


References
-
- Arnez, J.G., Moras, D. Structural and functional considerations of the aminoacylation reaction. Trends Biochem. Sci. 1997;22:211–216. - PubMed
-
- Balakrishnan, J. Symmetry scheme for amino acids codons. Phys. Rev. E. 2002;65:21912–21916. - PubMed
-
- Bedouelle, H. Recognition of tRNA-Tyr by tyrosyl-tRNA synthetase. Biochimie. 1990;72:589–598. - PubMed
-
- Berger, B., Leighton, T. Protein folding in the hydrophobic–hydrophilic HP model is NP-complete. J. Comput. Biol. 1998;5:27–40. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous