

Evolution of secretin family GPCR members in the metazoa
- PMID: 17166275
- PMCID: PMC1764030
- DOI: 10.1186/1471-2148-6-108
Evolution of secretin family GPCR members in the metazoa
Abstract
Background: Comparative approaches using protostome and deuterostome data have greatly contributed to understanding gene function and organismal complexity. The family 2 G-protein coupled receptors (GPCRs) are one of the largest and best studied hormone and neuropeptide receptor families. They are suggested to have arisen from a single ancestral gene via duplication events. Despite the recent identification of receptor members in protostome and early deuterostome genomes, relatively little is known about their function or origin during metazoan divergence. In this study a comprehensive description of family 2 GPCR evolution is given based on in silico and expression analyses of the invertebrate receptor genes.
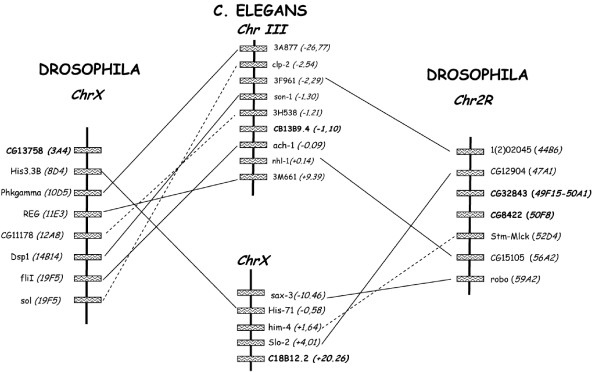
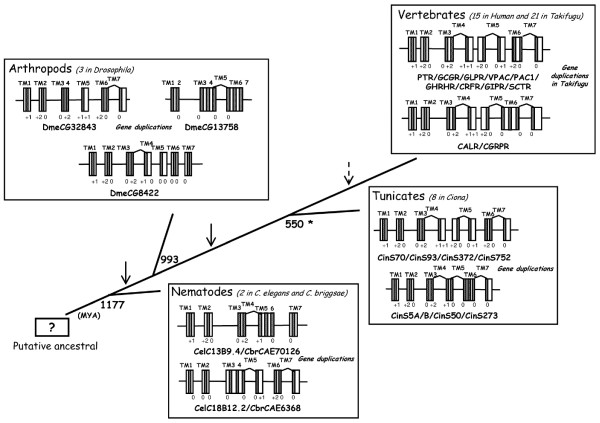
Results: Family 2 GPCR members were identified in the invertebrate genomes of the nematodes C. elegans and C. briggsae, the arthropods D. melanogaster and A. gambiae (mosquito) and in the tunicate C. intestinalis. This suggests that they are of ancient origin and have evolved through gene/genome duplication events. Sequence comparisons and phylogenetic analyses have demonstrated that the immediate gene environment, with regard to gene content, is conserved between the protostome and deuterostome receptor genomic regions. Also that the protostome genes are more like the deuterostome Corticotrophin Releasing Factor (CRF) and Calcitonin/Calcitonin Gene-Related Peptide (CAL/CGRP) receptors members than the other family 2 GPCR members. The evolution of family 2 GPCRs in deuterostomes is characterised by acquisition of new family members, with SCT (Secretin) receptors only present in tetrapods. Gene structure is characterised by an increase in intron number with organismal complexity with the exception of the vertebrate CAL/CGRP receptors.
Conclusion: The family 2 GPCR members provide a good example of gene duplication events occurring in tandem with increasing organismal complexity during metazoan evolution. The putative ancestral receptors are proposed to be more like the deuterostome CAL/CGRP and CRF receptors and this may be associated with their fundamental role in calcium regulation and the stress response, both of which are essential for survival.
Figures




Similar articles
-
The serendipitous origin of chordate secretin peptide family members.BMC Evol Biol. 2010 May 6;10:135. doi: 10.1186/1471-2148-10-135. BMC Evol Biol. 2010. PMID: 20459630 Free PMC article.
-
Nematode and arthropod genomes provide new insights into the evolution of class 2 B1 GPCRs.PLoS One. 2014 Mar 20;9(3):e92220. doi: 10.1371/journal.pone.0092220. eCollection 2014. PLoS One. 2014. PMID: 24651821 Free PMC article.
-
Prevertebrate Local Gene Duplication Facilitated Expansion of the Neuropeptide GPCR Superfamily.Mol Biol Evol. 2015 Nov;32(11):2803-17. doi: 10.1093/molbev/msv179. Epub 2015 Sep 3. Mol Biol Evol. 2015. PMID: 26337547
-
New insights into the evolution of vertebrate CRH (corticotropin-releasing hormone) and invertebrate DH44 (diuretic hormone 44) receptors in metazoans.Gen Comp Endocrinol. 2014 Dec 1;209:162-70. doi: 10.1016/j.ygcen.2014.09.004. Epub 2014 Sep 16. Gen Comp Endocrinol. 2014. PMID: 25230393 Review.
-
Intermedin, a novel calcitonin family peptide that exists in teleosts as well as in mammals: a comparison with other calcitonin/intermedin family peptides in vertebrates.Peptides. 2004 Oct;25(10):1633-42. doi: 10.1016/j.peptides.2004.05.021. Peptides. 2004. PMID: 15476930 Review.
Cited by
-
Prediction of G Protein-Coupled Receptors with SVM-Prot Features and Random Forest.Scientifica (Cairo). 2016;2016:8309253. doi: 10.1155/2016/8309253. Epub 2016 Jul 27. Scientifica (Cairo). 2016. PMID: 27529053 Free PMC article.
-
Systematic study on G-protein couple receptor prototypes: did they really evolve from prokaryotic genes?IET Syst Biol. 2014 Aug;8(4):154-61. doi: 10.1049/iet-syb.2013.0037. IET Syst Biol. 2014. PMID: 25075528 Free PMC article.
-
Interdependence between SEB-3 receptor and NLP-49 peptides shifts across predator-induced defensive behavioral modes in Caenorhabditis elegans.Elife. 2025 Mar 31;13:RP98262. doi: 10.7554/eLife.98262. Elife. 2025. PMID: 40163376 Free PMC article.
-
Evolution of parathyroid hormone receptor family and their ligands in vertebrate.Front Endocrinol (Lausanne). 2015 Mar 10;6:28. doi: 10.3389/fendo.2015.00028. eCollection 2015. Front Endocrinol (Lausanne). 2015. PMID: 25806022 Free PMC article. Review.
-
The serendipitous origin of chordate secretin peptide family members.BMC Evol Biol. 2010 May 6;10:135. doi: 10.1186/1471-2148-10-135. BMC Evol Biol. 2010. PMID: 20459630 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
