

Modeling the evolution of protein domain architectures using maximum parsimony
- PMID: 17166515
- PMCID: PMC1858635
- DOI: 10.1016/j.jmb.2006.11.017
Modeling the evolution of protein domain architectures using maximum parsimony
Abstract
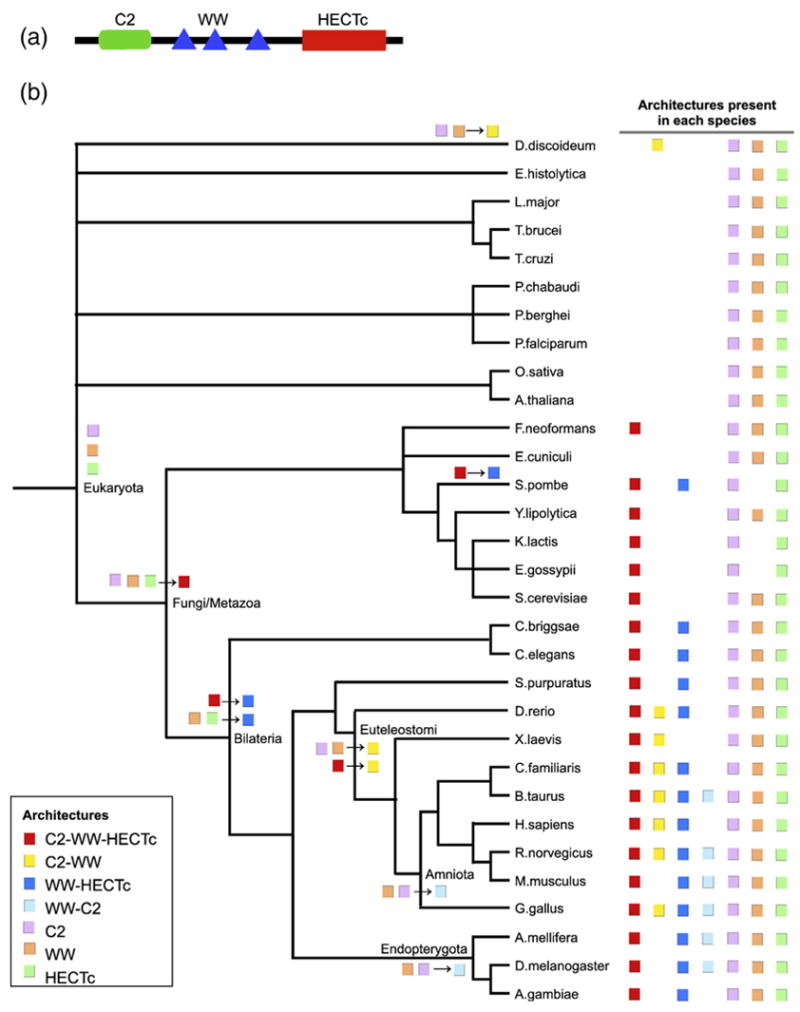
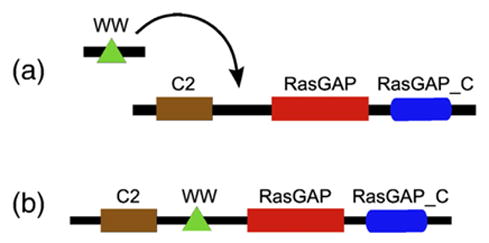
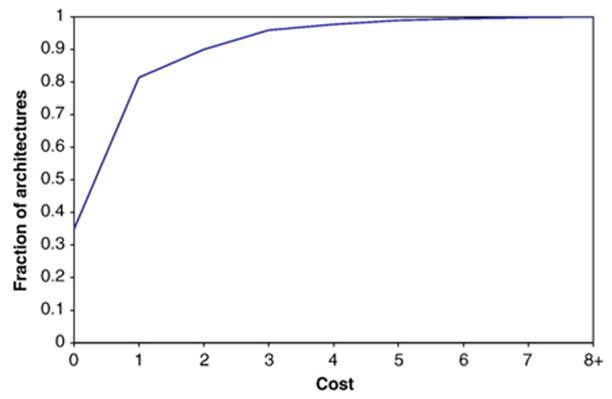
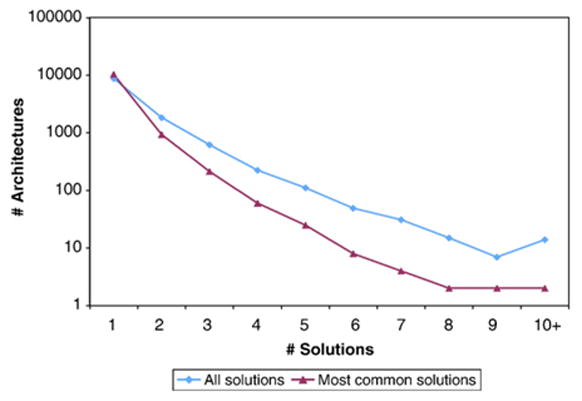
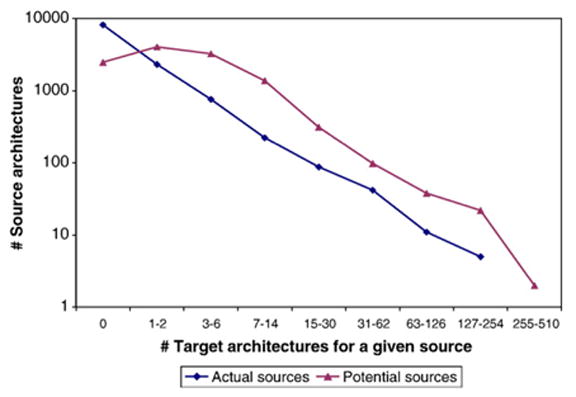
Domains are basic evolutionary units of proteins and most proteins have more than one domain. Advances in domain modeling and collection are making it possible to annotate a large fraction of known protein sequences by a linear ordering of their domains, yielding their architecture. Protein domain architectures link evolutionarily related proteins and underscore their shared functions. Here, we attempt to better understand this association by identifying the evolutionary pathways by which extant architectures may have evolved. We propose a model of evolution in which architectures arise through rearrangements of inferred precursor architectures and acquisition of new domains. These pathways are ranked using a parsimony principle, whereby scenarios requiring the fewest number of independent recombination events, namely fission and fusion operations, are assumed to be more likely. Using a data set of domain architectures present in 159 proteomes that represent all three major branches of the tree of life allows us to estimate the history of over 85% of all architectures in the sequence database. We find that the distribution of rearrangement classes is robust with respect to alternative parsimony rules for inferring the presence of precursor architectures in ancestral species. Analyzing the most parsimonious pathways, we find 87% of architectures to gain complexity over time through simple changes, among which fusion events account for 5.6 times as many architectures as fission. Our results may be used to compute domain architecture similarities, for example, based on the number of historical recombination events separating them. Domain architecture "neighbors" identified in this way may lead to new insights about the evolution of protein function.
Figures
References
-
- Bork P. Mobile modules and motifs. Curr Opin Struct Biol. 1992;2:413–421.
-
- Murzin AG, Brenner SE, Hubbard T, Chothia C. SCOP: a structural classification of proteins database for the investigation of sequences and structures. J Mol Biol. 1995;247:536–540. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
