

Phylogeny of the order Choreotrichida (Ciliophora, Spirotricha, Oligotrichea) as inferred from morphology, ultrastructure, ontogenesis, and SSrRNA gene sequences
- PMID: 17166704
- PMCID: PMC2850171
- DOI: 10.1016/j.ejop.2006.10.001
Phylogeny of the order Choreotrichida (Ciliophora, Spirotricha, Oligotrichea) as inferred from morphology, ultrastructure, ontogenesis, and SSrRNA gene sequences
Abstract
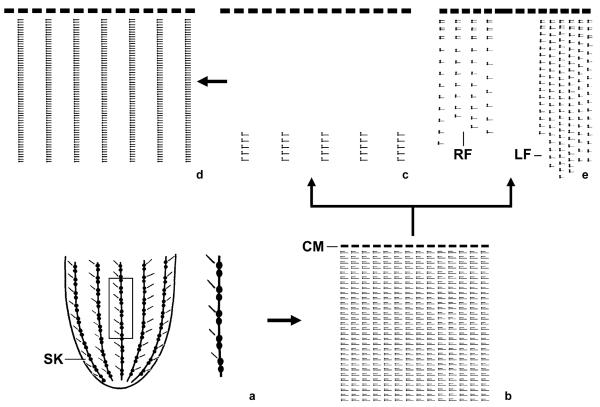
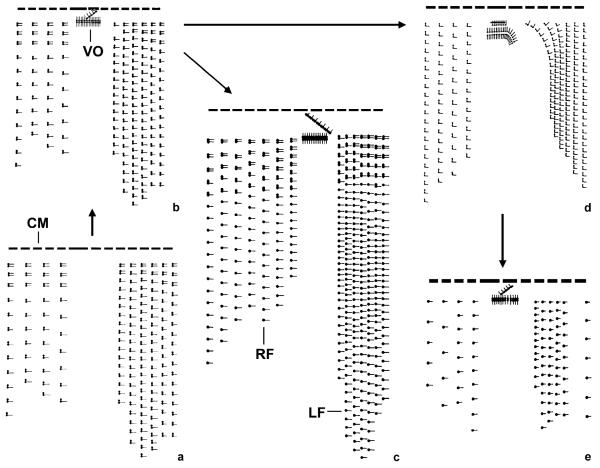
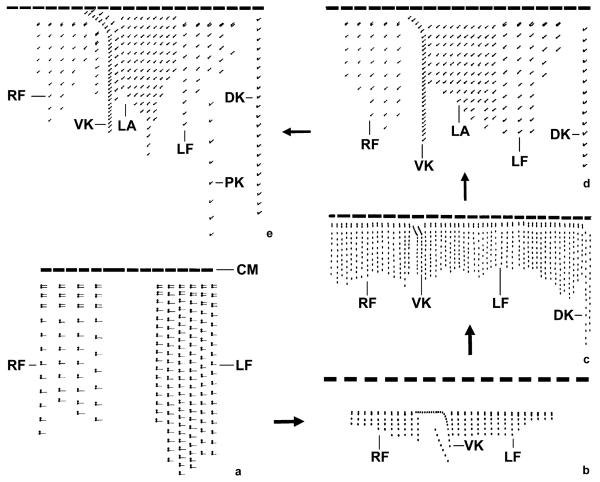
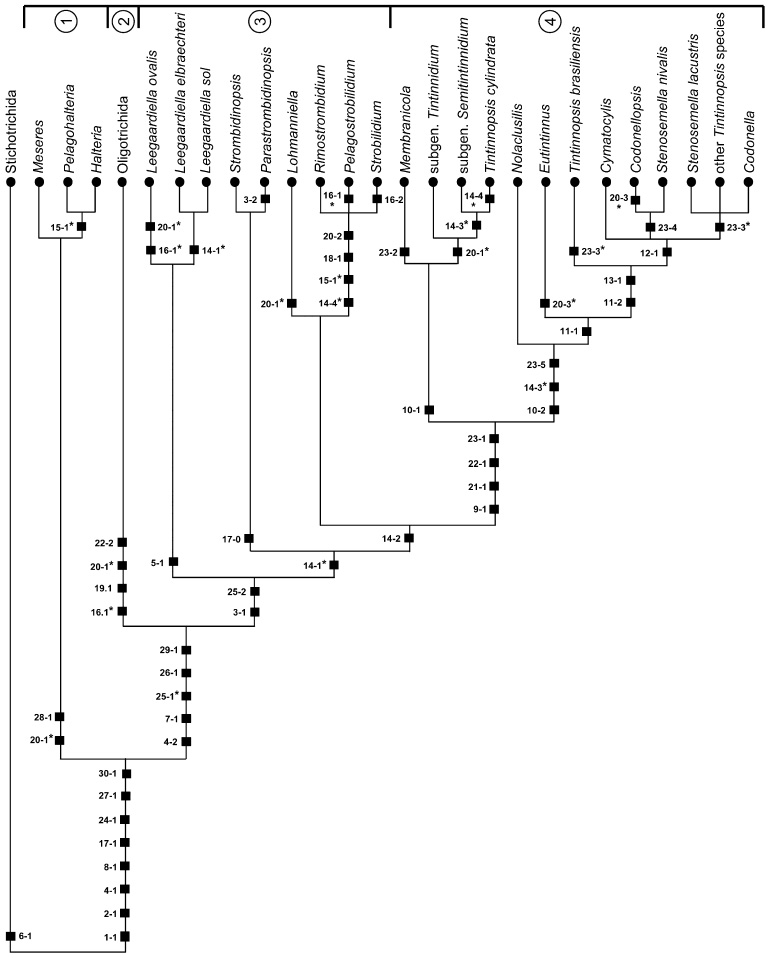
The phylogeny within the order Choreotrichida is reconstructed using (i) morphologic, ontogenetic, and ultrastructural evidence for the cladistic approach and (ii) the small subunit ribosomal RNA (SSrRNA) gene sequences, including the new sequence of Rimostrombidium lacustris. The morphologic cladograms and the gene trees converge rather well for the Choreotrichida, demonstrating that hyaline and agglutinated loricae do not characterize distinct lineages, i.e., both lorica types can be associated with the most highly developed ciliary pattern. The position of Rimostrombidium lacustris within the family Strobilidiidae is corroborated by the genealogical analyses. The diagnosis of the genus Tintinnidium is improved, adding cytological features, and the genus is divided into two subgenera based on the structure of the somatic kineties. The diagnosis of the family Lohmanniellidae and the genus Lohmanniella are improved, and Rimostrombidium glacicolum Petz, Song and Wilbert, 1995 is affiliated.
Figures
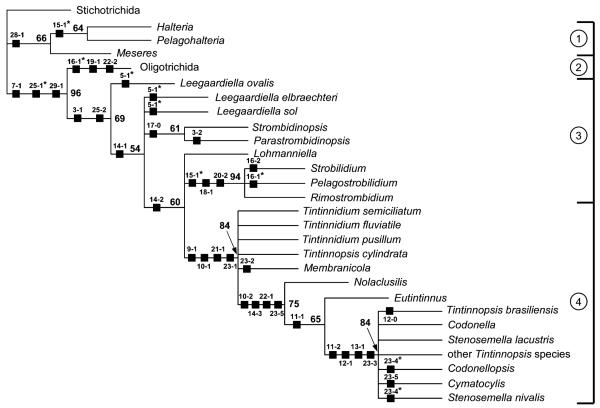
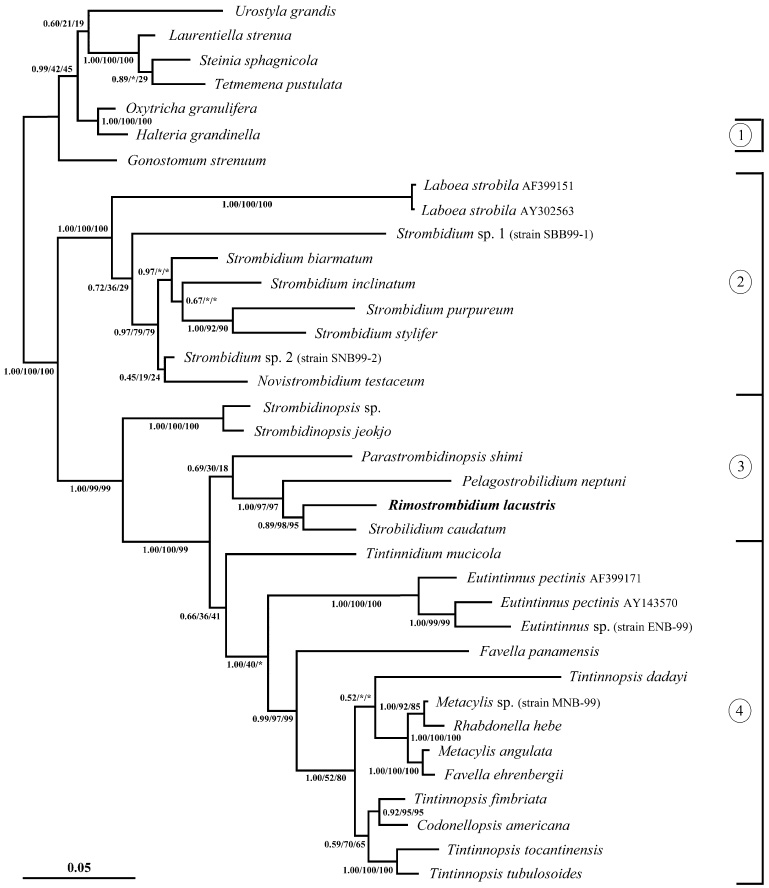
References
-
- Agatha S. Morphology and ontogenesis of Novistrombidium apsheronicum nov. comb. and Strombidium arenicola (Protozoa, Ciliophora): a comparative light microscopical and SEM study. Eur. J. Protistol. 2003a;39:245–266.
-
- Agatha S. Redescription of Strombidinopsis minima (Gruber, 1884) Lynn et al., 1991 (Protozoa, Ciliophora), with notes on its ontogenesis and distribution. Eur. J. Protistol. 2003b;39:233–244.
-
- Agatha S. New observations on the tontoniid ciliate Spirotontonia grandis (Suzuki and Han, 2000) Agatha, 2004 (Ciliophora, Oligotrichida, Tontoniidae); comparison with the similar Laboea strobila. Eur. J. Protistol. 2004c;40:295–301.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
