

Basolateral entry and release of Crimean-Congo hemorrhagic fever virus in polarized MDCK-1 cells
- PMID: 17166898
- PMCID: PMC1865934
- DOI: 10.1128/JVI.02070-06
Basolateral entry and release of Crimean-Congo hemorrhagic fever virus in polarized MDCK-1 cells
Abstract
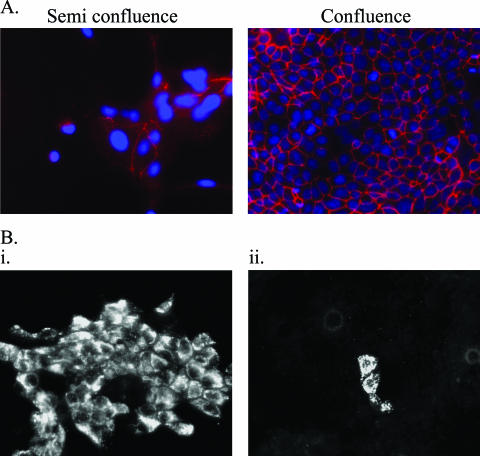
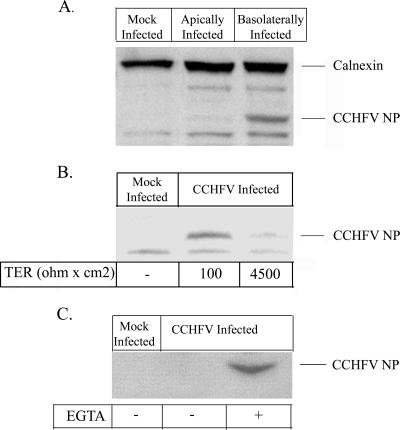
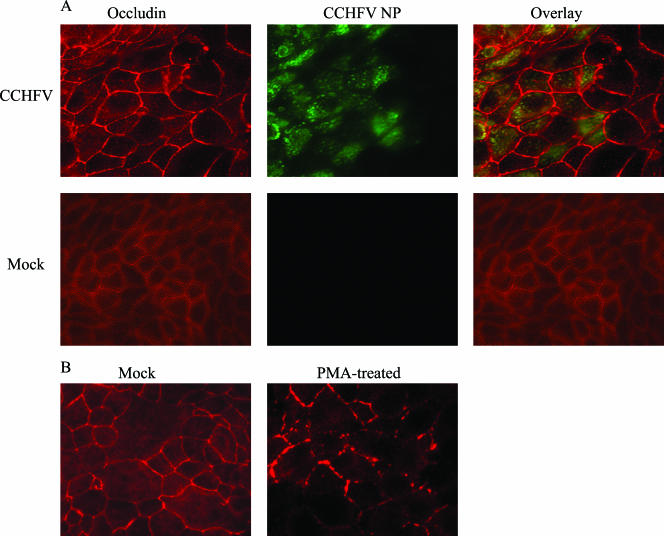
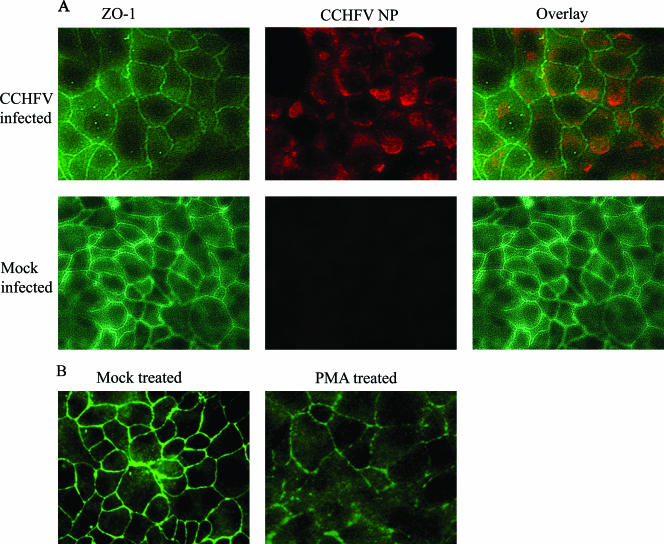
Crimean-Congo hemorrhagic fever virus (CCHFV) is an etiological agent of a disease with mortality rates in patients averaging 30%. The disease is characterized by fever, myalgia, and hemorrhage. Mechanisms underlying the hemorrhage have to our knowledge not been elucidated for CCHFV. Possibly, a direct or indirect viral effect on tight junctions (TJ) could cause the hemorrhage observed in patients, as TJ play a crucial role in vascular homeostasis and can cause leakage upon deregulation. Moreover, there is no knowledge regarding the site of entry and release of CCHFV in polarized epithelial cells. Such cells represent a barrier to virus dissemination within the host, and as a site of viral entry and release, they could play a key role in further spread. For the first time, we have shown preferential basolateral entry of CCHFV in Madin-Darby canine kidney 1 (MDCK-1) epithelial cells. Furthermore, we demonstrated basolateral release of CCHFV in polarized epithelial cells. Interestingly, by measuring transepithelial electrical resistance, we found no effect of CCHFV replication on the function of TJ in this study. Neither did we observe any difference in the localization of the TJ proteins ZO-1 and occludin in CCHFV-infected cells compared to mock-infected cells.
Figures
References
-
- Andersson, I., M. Simon, A. Lundkvist, M. Nilsson, A. Holmstrom, F. Elgh, and A. Mirazimi. 2004. Role of actin filaments in targeting of Crimean Congo hemorrhagic fever virus nucleocapsid protein to perinuclear regions of mammalian cells. J. Med. Virol. 72:83-93. - PubMed
-
- Bazzoni, G., and E. Dejana. 2004. Endothelial cell-to-cell junctions: molecular organization and role in vascular homeostasis. Physiol. Rev. 84:869-901. - PubMed
-
- Bertolotti-Ciarlet, A., J. Smith, K. Strecker, J. Paragas, L. A. Altamura, J. M. McFalls, N. Frias-Staheli, A. Garcia-Sastre, C. S. Schmaljohn, and R. W. Doms. 2005. Cellular localization and antigenic characterization of Crimean-Congo hemorrhagic fever virus glycoproteins. J. Virol. 79:6152-6161. - PMC - PubMed
-
- Bosch, I., K. Xhaja, L. Estevez, G. Raines, H. Melichar, R. V. Warke, M. V. Fournier, F. A. Ennis, and A. L. Rothman. 2002. Increased production of interleukin-8 in primary human monocytes and in human epithelial and endothelial cell lines after dengue virus challenge. J. Virol. 76:5588-5597. - PMC - PubMed
-
- Bray, M., and T. W. Geisbert. 2005. Ebola virus: the role of macrophages and dendritic cells in the pathogenesis of Ebola hemorrhagic fever. Int. J. Biochem. Cell Biol. 37:1560-1566. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
