

The linear computational algorithm of cerebellar Purkinje cells
- PMID: 17167077
- PMCID: PMC6674952
- DOI: 10.1523/JNEUROSCI.4507-05.2006
The linear computational algorithm of cerebellar Purkinje cells
Abstract
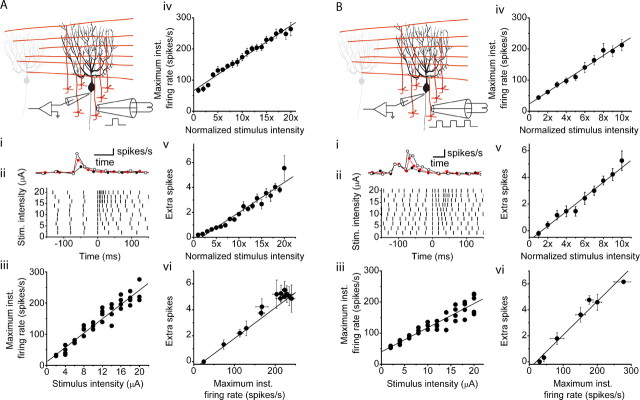
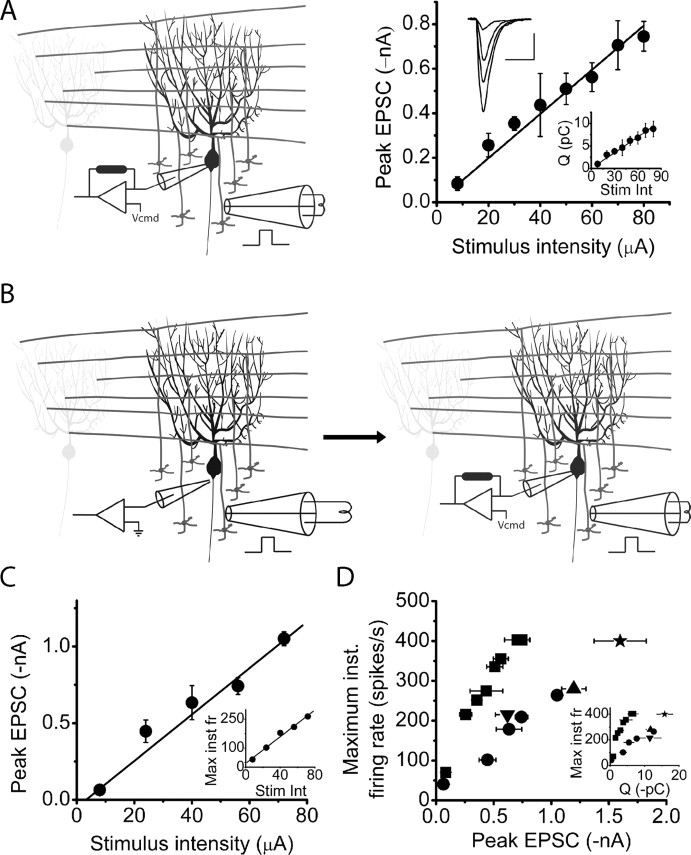
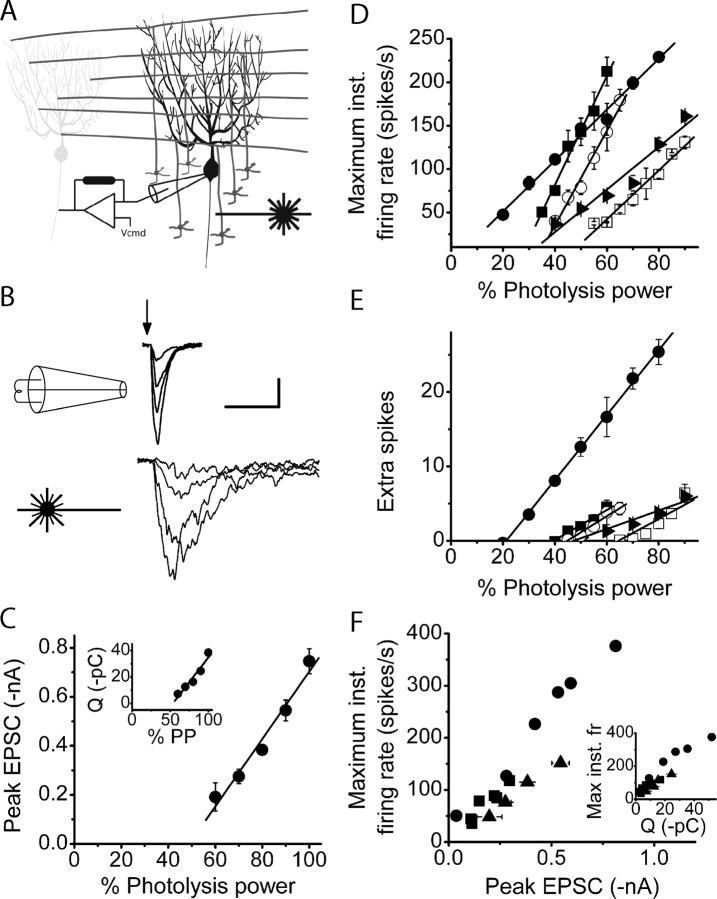
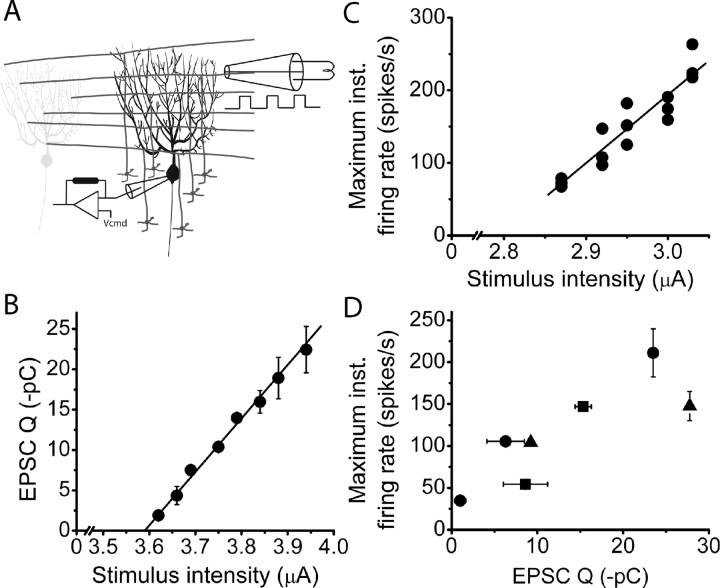
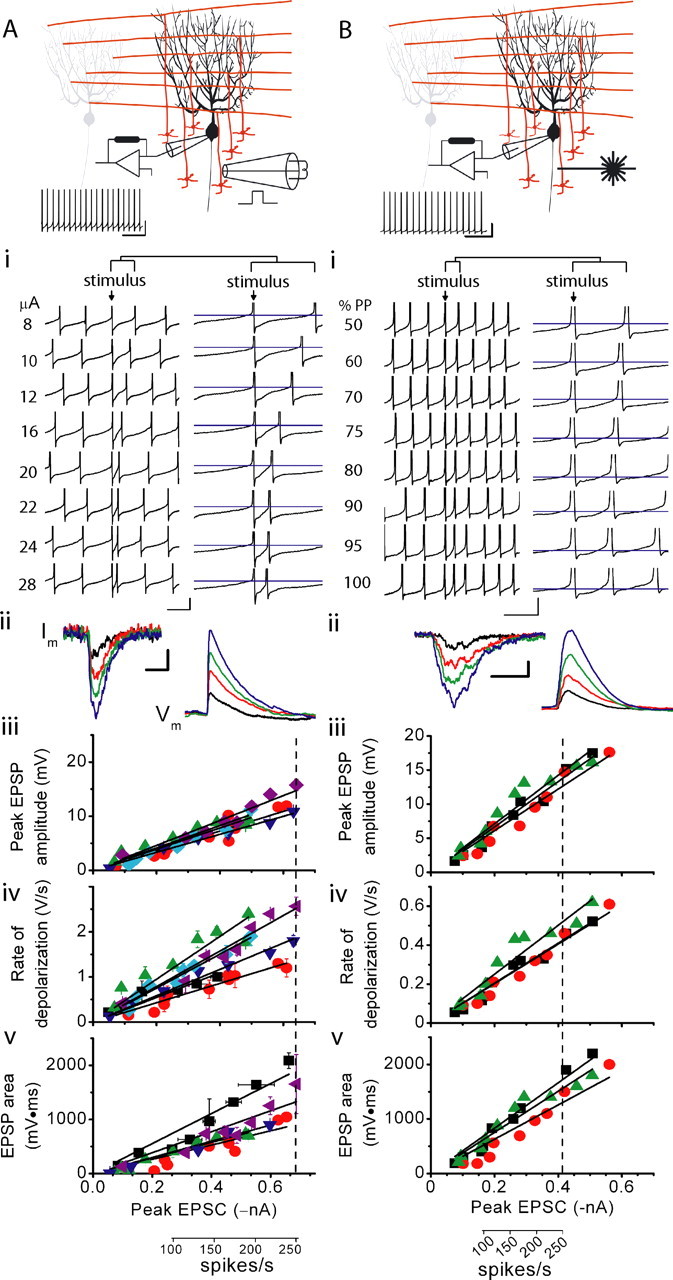
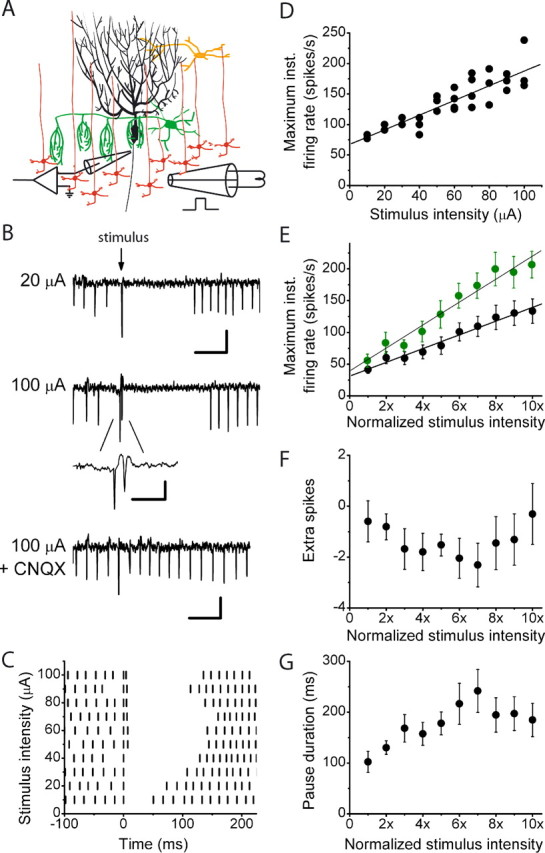
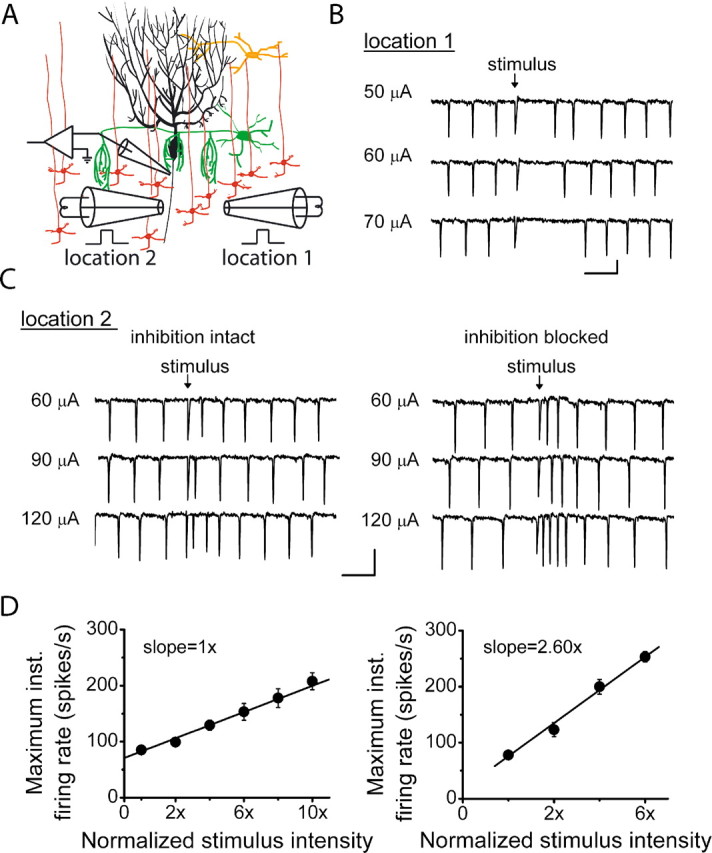
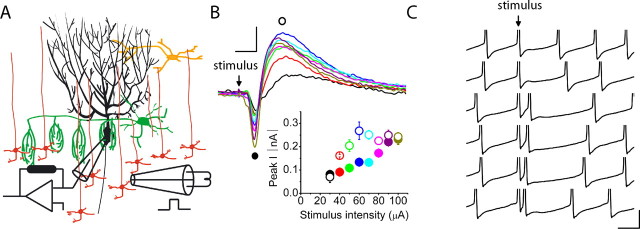
The orchestration of simple motor tasks by the cerebellum results in coordinated movement and the maintenance of balance. The cerebellum integrates sensory and cortical information to generate the signals required for the coordinated execution of simple motor tasks. These signals originate in the firing rate of Purkinje cells, each of which integrates sensory and cortical information conveyed by granule cell synaptic inputs. Given the importance of the granule cell input-Purkinje cell output function for cerebellar computation, this algorithm was determined. Using several stimulation paradigms, including those that mimicked patterns of granule cell activity similar to those observed in vivo, we quantified the poststimulus maximum firing rate and number of extra spikes in response to granule cell synaptic input. Both of these parameters linearly encoded the strength of synaptic input when inhibitory synaptic transmission was blocked. This linear algorithm was independent of the location or temporal pattern of synaptic input. With inhibitory synaptic transmission intact, the maximum firing rate, but not the number of extra spikes, encoded the strength of granule cell synaptic input. Furthermore, the maximum firing rate of Purkinje cells linearly encoded the strength of synaptic input whether or not the activation of granule cells resulted in a pause in Purkinje cell firing. On the basis of the data presented, we propose that Purkinje cells encode the strength of granule cell synaptic input in their maximum firing rate with a linear algorithm.
Figures
References
-
- Albus JS. A theory of cerebellar function. Math Biosci. 1971;10:25–61.
-
- Allen GI, Azzena GB, Ohno T. Somatotopically organized inputs from fore- and hindlimb areas of sensorimotor cortex to cerebellar Purkyne cells. Exp Brain Res. 1974;20:255–272. - PubMed
-
- Bower JM, Woolston DC. Congruence of spatial organization of tactile projections to granule cell and Purkinje cell layers of cerebellar hemispheres of the albino rat: vertical organization of cerebellar cortex. J Neurophysiol. 1983;49:745–766. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources