

Review
doi: 10.1021/jo0614240.
Rings, radicals, and regeneration: the early years of a bioorganic laboratory
Affiliations
- PMID: 17168571
- PMCID: PMC2519235
- DOI: 10.1021/jo0614240
Item in Clipboard
Review
Rings, radicals, and regeneration: the early years of a bioorganic laboratory
J Org Chem.
.
Abstract
This Perspective provides an overview of the progress in two of the original programs in my research group focused on the biosynthesis of the antibiotics nisin, lacticin 481, fosfomycin, and bialaphos. The path from start-up funds to tenure and beyond offers insights into the opportunities realized and missed along the road.
Figures
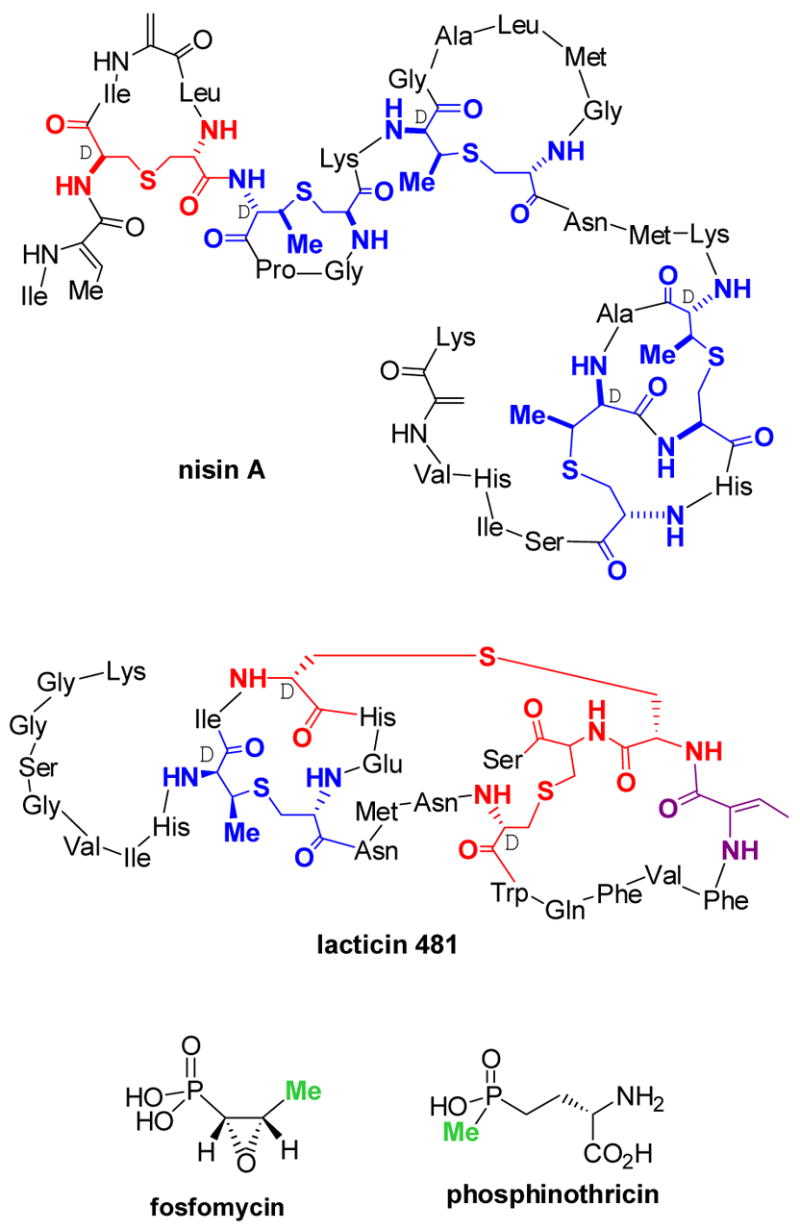
Structures of the lantibiotics nisin A and lacticin 481, the phosphonate antibiotic fosfomycin, and the phosphinate phosphinothricin. Lanthionine (Lan) residues in the lantibiotics are shown in red and methyllanthionine (MeLan) residues in blue. The two methyl groups in fosfomycin and phosphinothricin that are considered to be derived from methylcobalamin are highlighted in green.
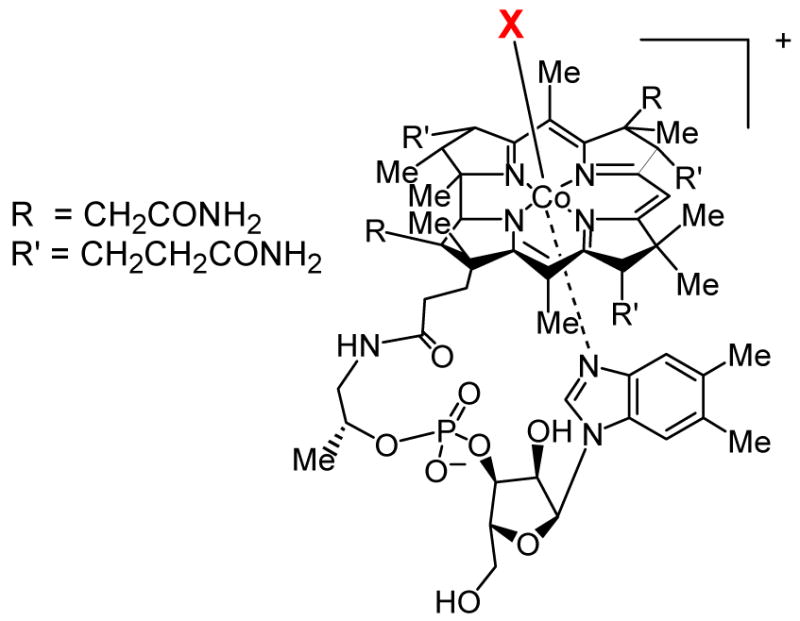
Structures of vitamin B12 (X = cyanide), and the natural cofactors methylcobalamin (X = CH3) and adenosylcobalamin (X = 5′-deoxyadenosine).
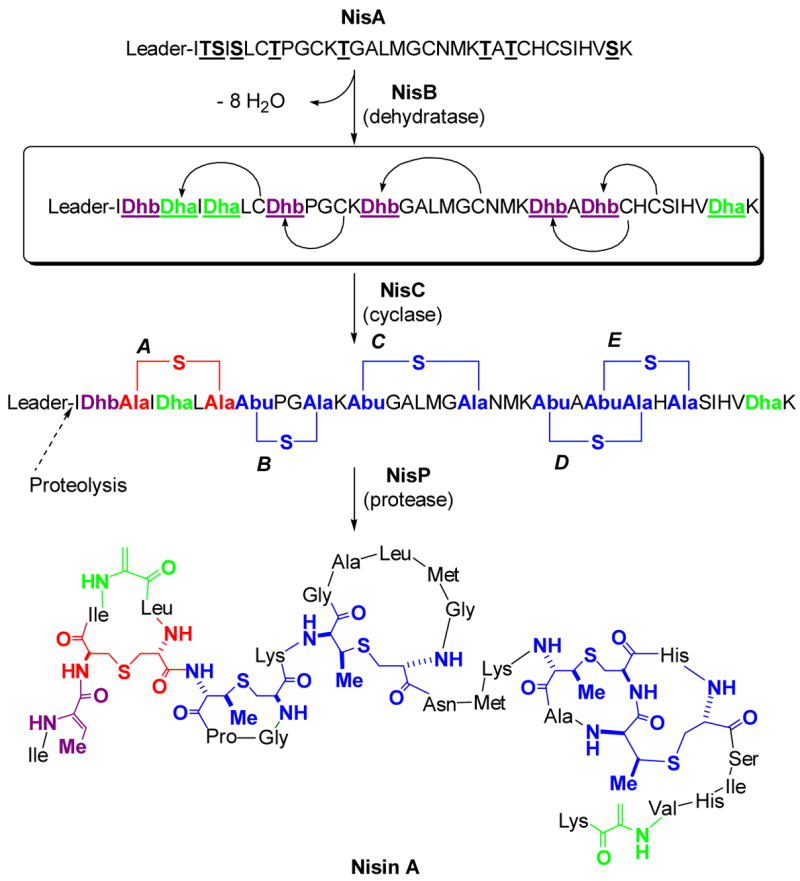
The post-translational maturation process of nisin as an example of a class I lantibiotic. The prepeptide NisA is ribosomally synthesized, followed by NisB catalyzed dehydration of underlined Ser and Thr residues of NisA. NisC catalyzes the conjugate addition of Cys residues in a regio- and stereoselective manner to five of the Dha (green) and Dhb (magenta) residues to generate five cyclic thioethers: one lanthionine (red) and four methyllanthionines (blue). After dehydration/cyclization is complete, the unmodified leader peptide is proteolytically removed by the protease NisP. The sequence of the leader peptide is MSTKDFNLDLVSVSKKDSGASPR. Abu, S-2-aminobutyric acid.
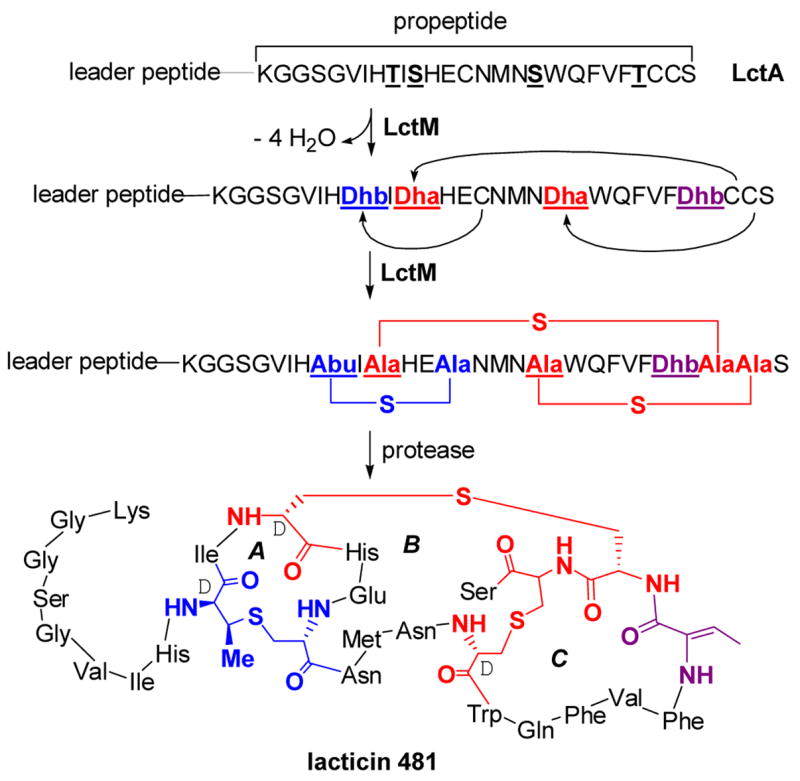
The post-translational maturation process of lacticin 481, a class II lantibiotic. LctM catalyzes the anti elimination of water from the underlined Ser and Thr residues in the propeptide region of LctA to generate Dha and Dhb residues. LctM also catalyzes the conjugate addition of three Cys residues in a regioselective manner to three of the dehydrated residues to generate three cyclic thioethers, one methyllanthionine (blue) and two lanthionines (red). The leader peptide is proteolytically removed by the N-terminal protease domain of LctT that also excretes the final product. The sequence of the leader peptide is MKEQNSFNLLQEVTESELDLILGA. Abu, S-2-aminobutyric acid.
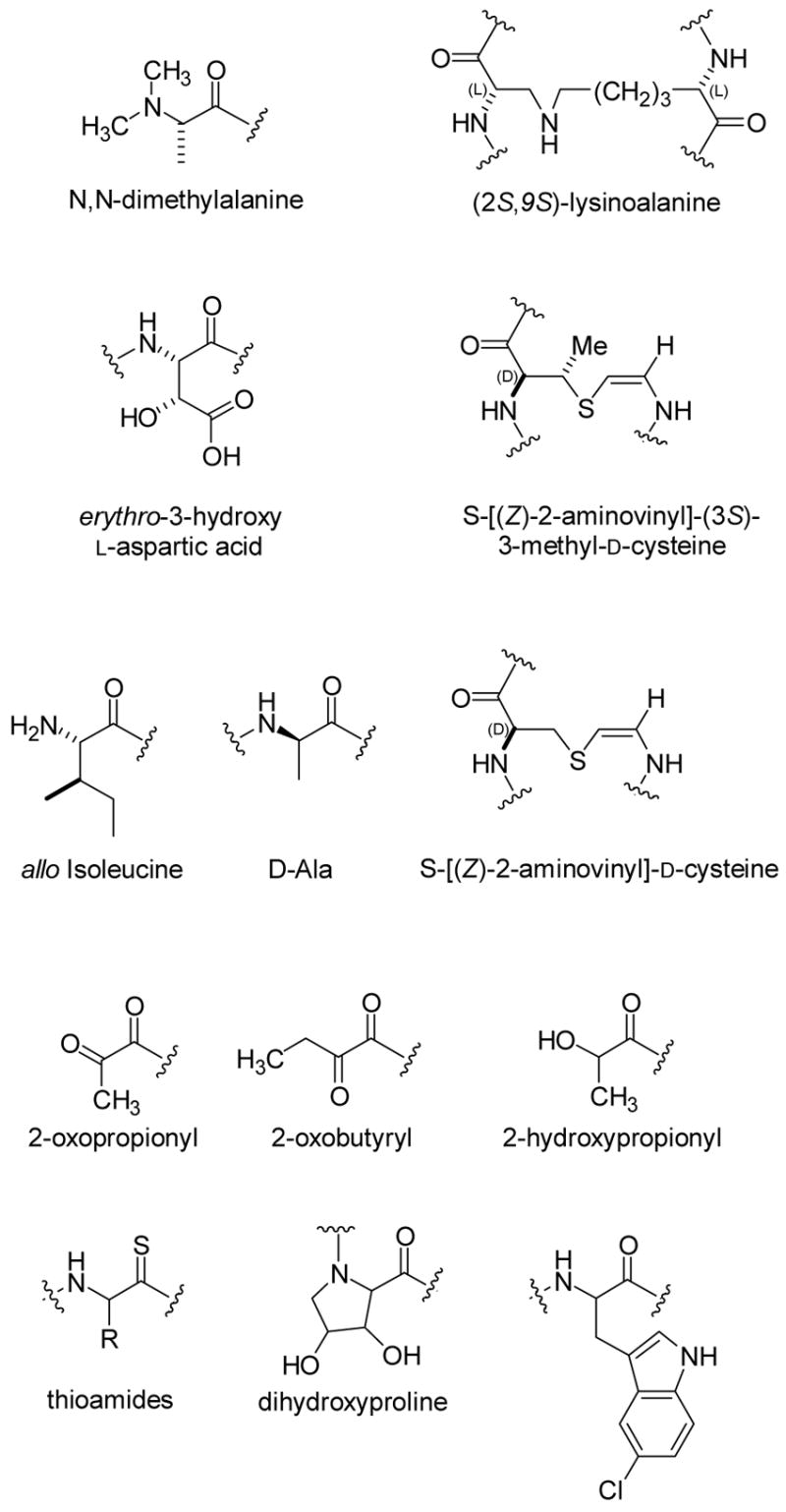
Various post-translational modifications of proteinogenic amino acids found in lantibiotic family members. The stereochemistry of the dihydroxyproline is currently unknown. Thioviramide contains an aminovinylcysteine suggesting it may be a lantibiotic, but it has not yet been shown to be gene encoded. It also contains the thioamides shown in the figure.
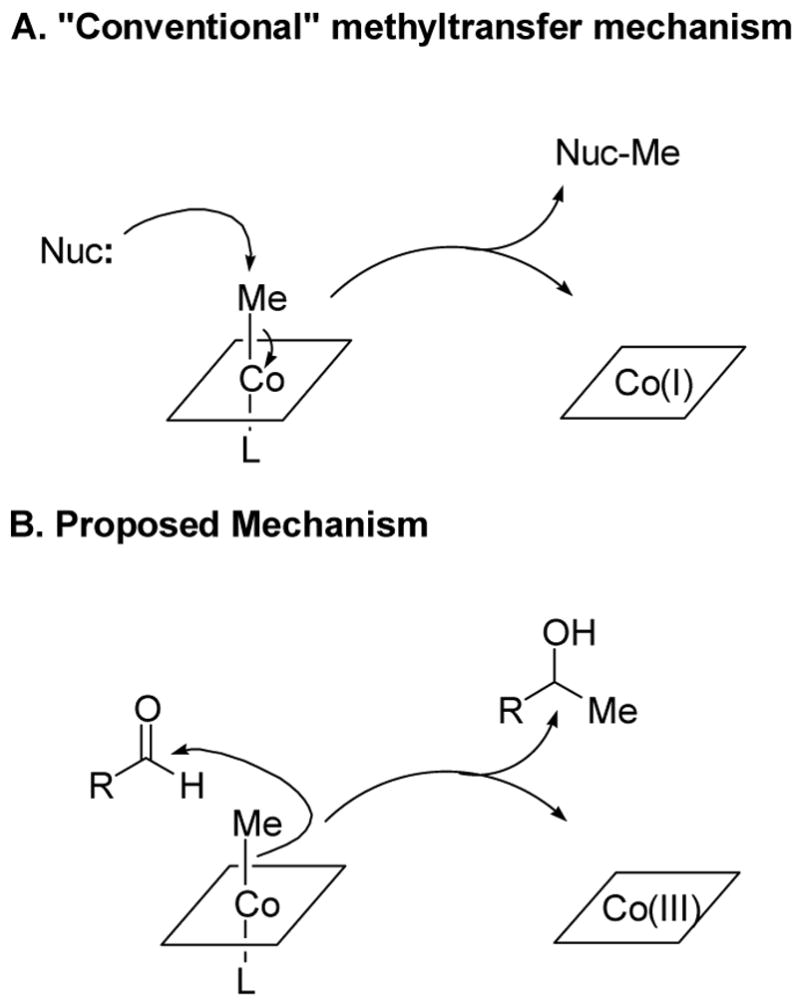
Conventional and unusual mechanisms, of B12-dependent methyl transfer. The corrin ligand ring system (Figure 2) is schematically represented by a square. L may be the benzimidazole ligand of B12 or an amino acid ligand from the protein.
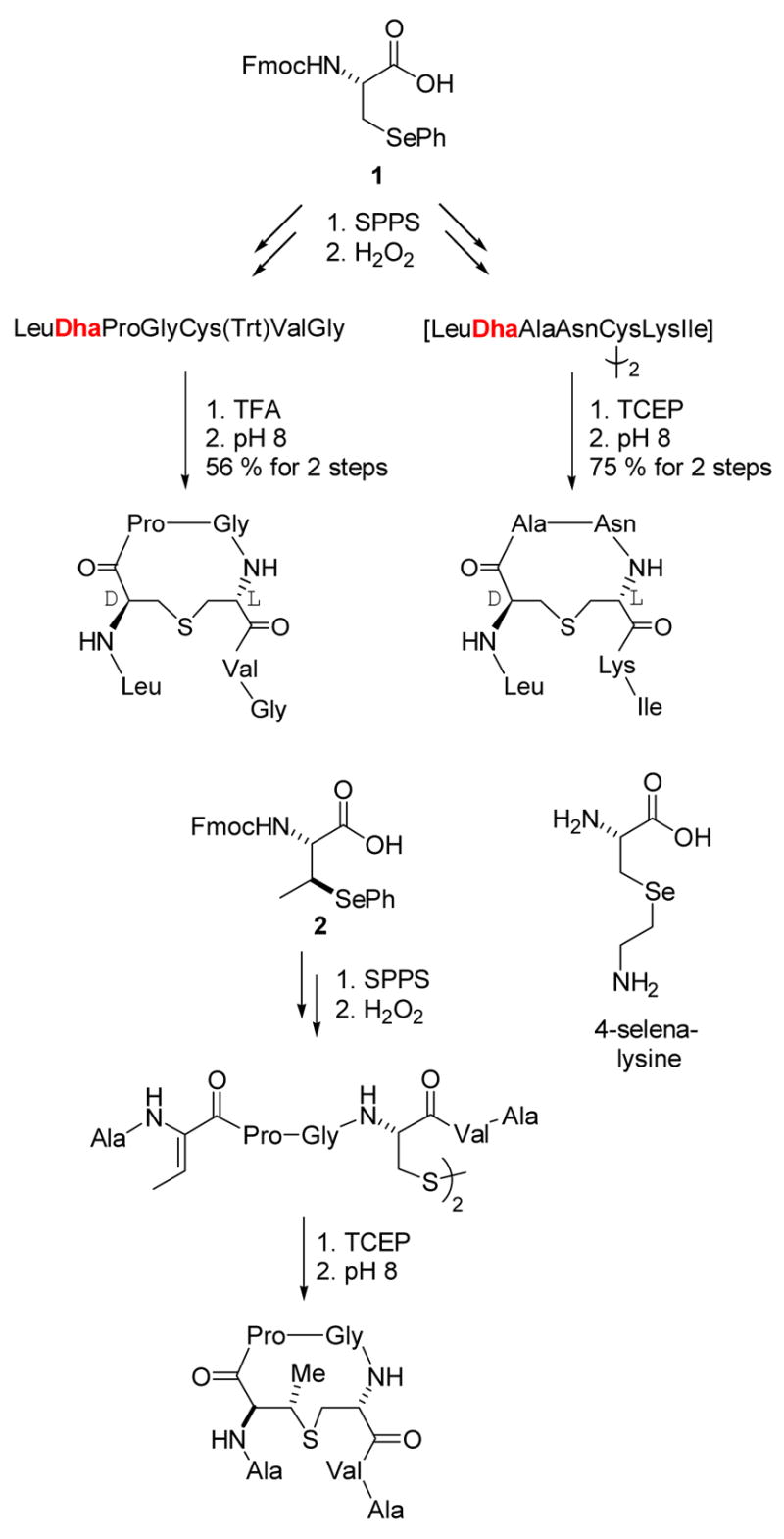
Preparation of dehydropeptides by oxidative elimination.
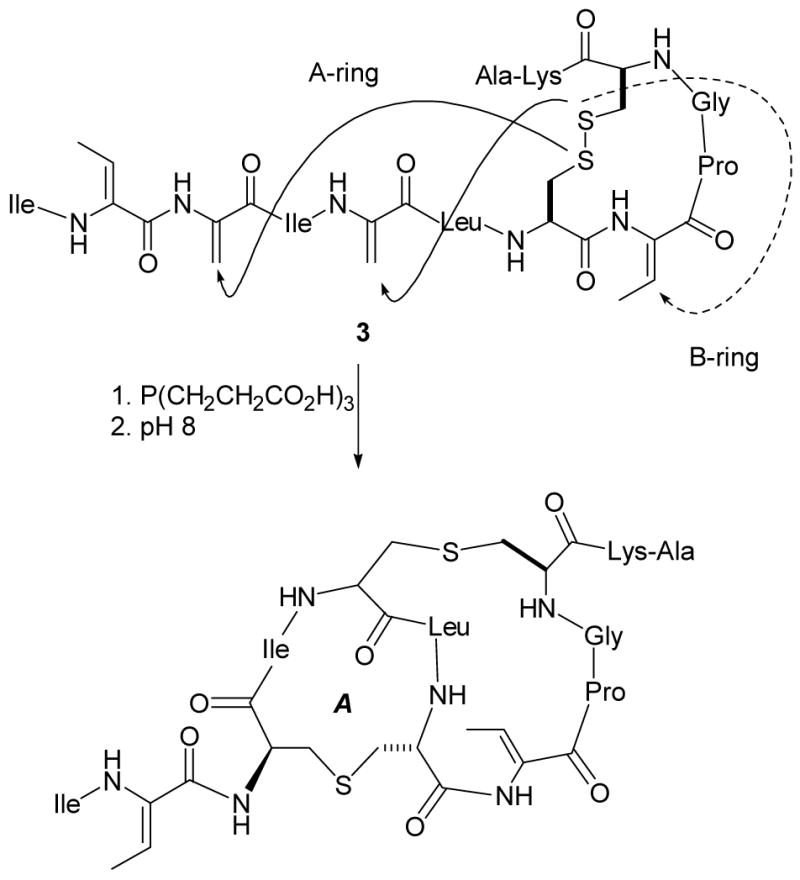
Attempted biomimetic synthesis of the A and B-rings of nisin.
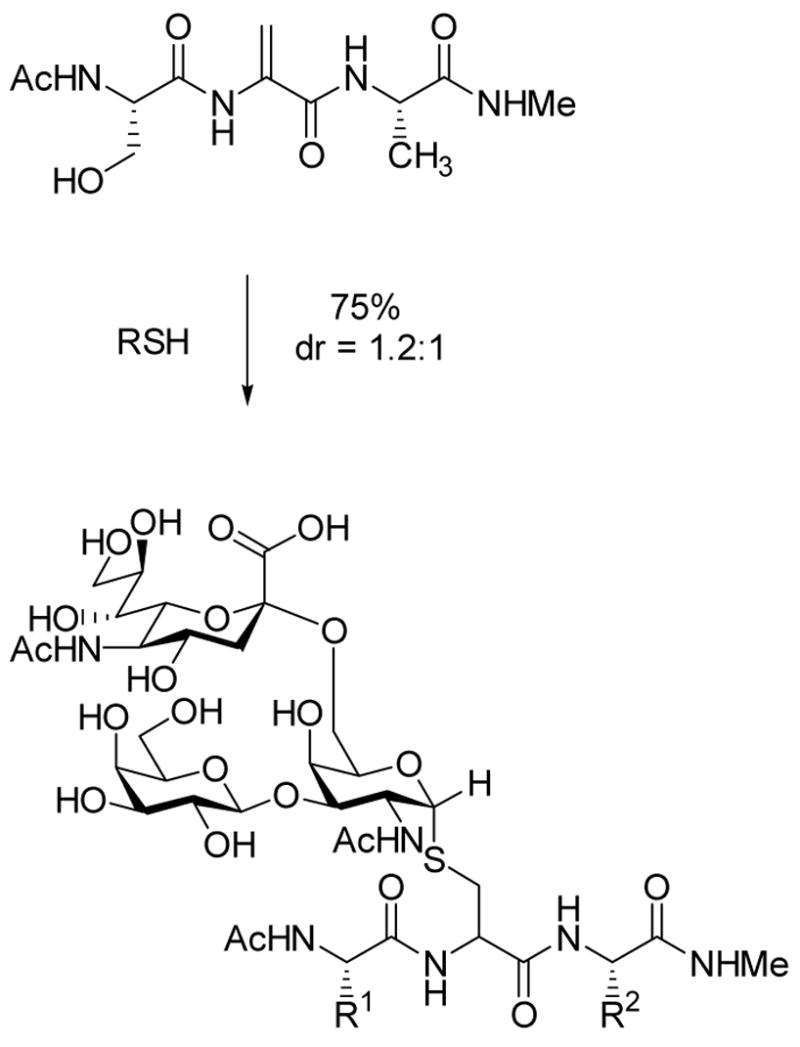
Conjugate addition of external thiol nucleophiles to Dha.
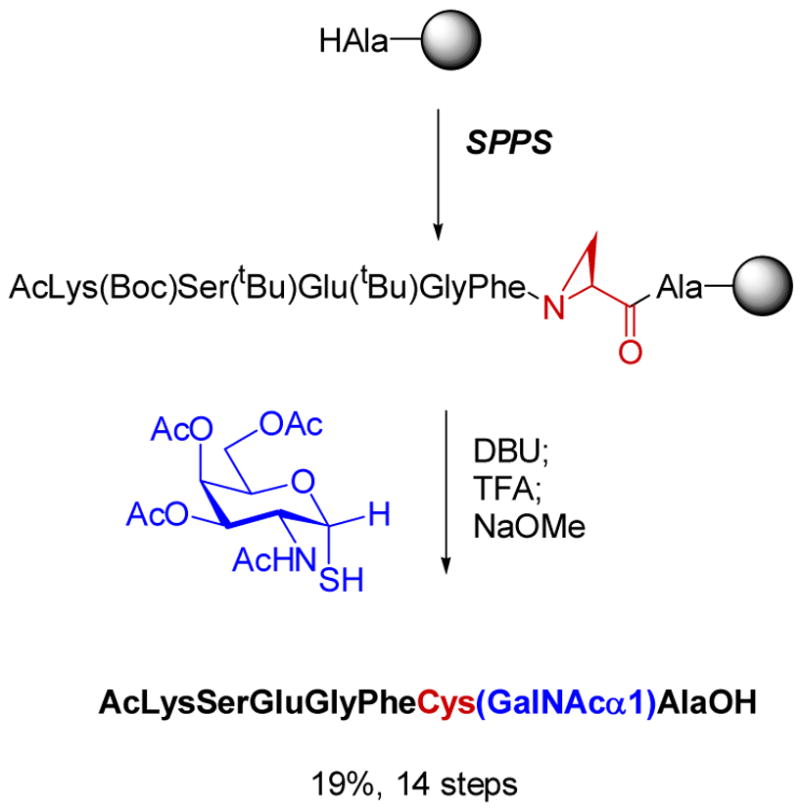
Stereoselective synthesis of thioglycopeptides.,
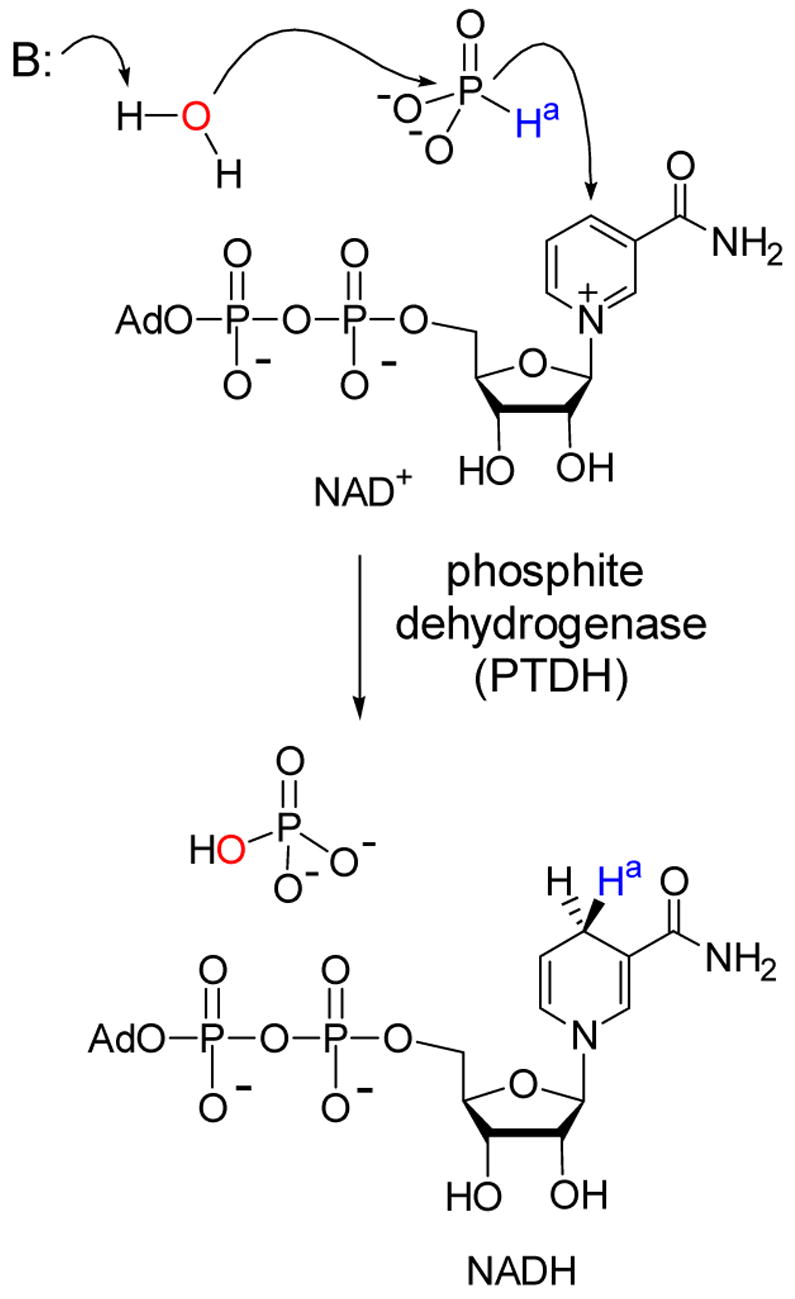
NAD+-dependent oxidation of phosphite by PTDH.
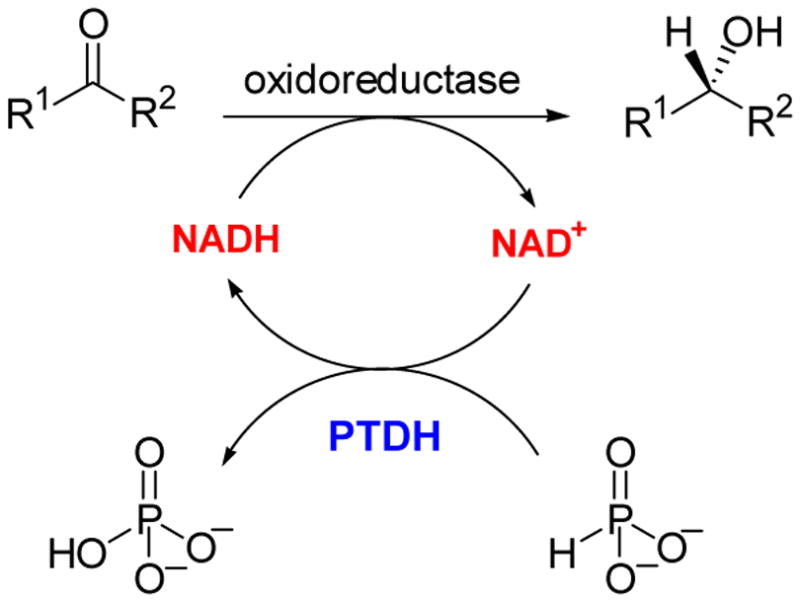
Use of PTDH for cofactor regeneration.
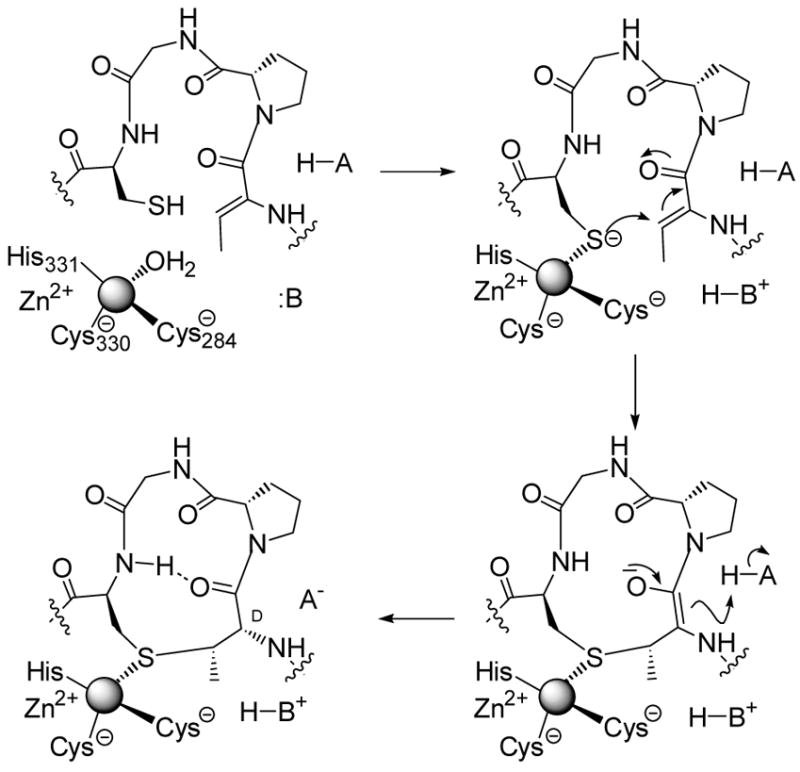
Proposed mechanism of enzymatic Lan and MeLan formation.
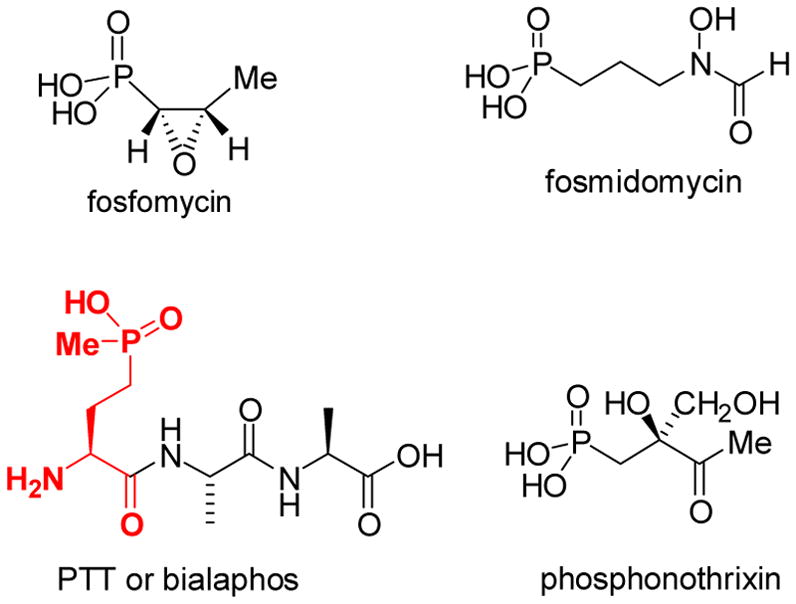
Several members of the phosphonate/phosphinate class of natural products. The phosphinothricin moiety in bialaphos is highlighted in red.
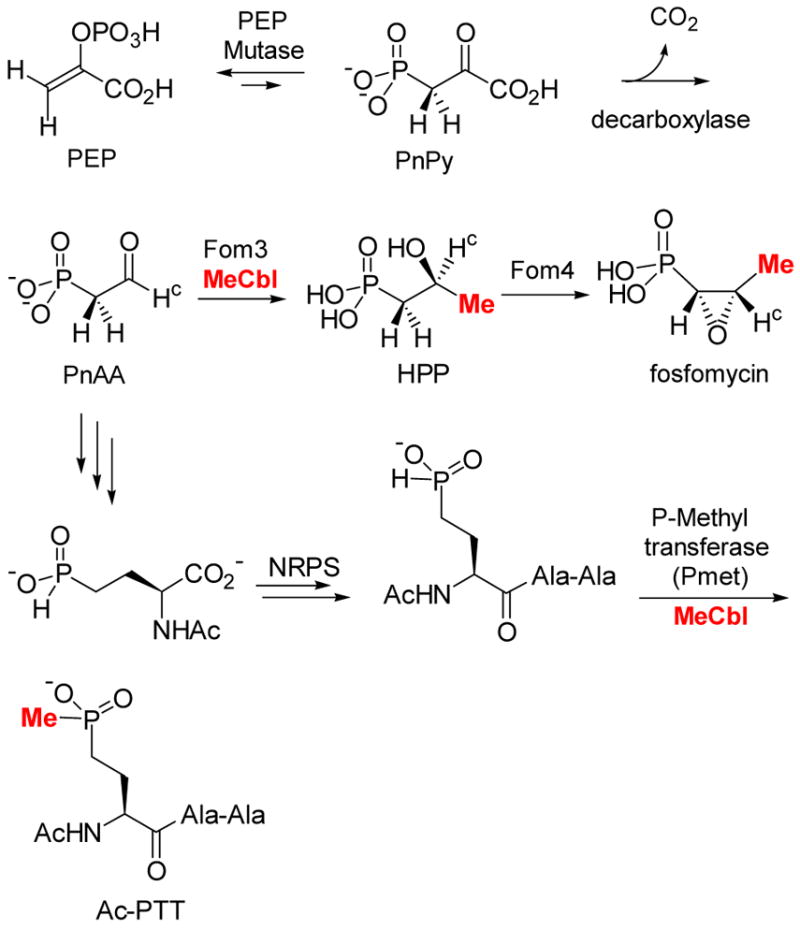
Proposed biosynthetic pathways of fosfomycin and bialaphos.
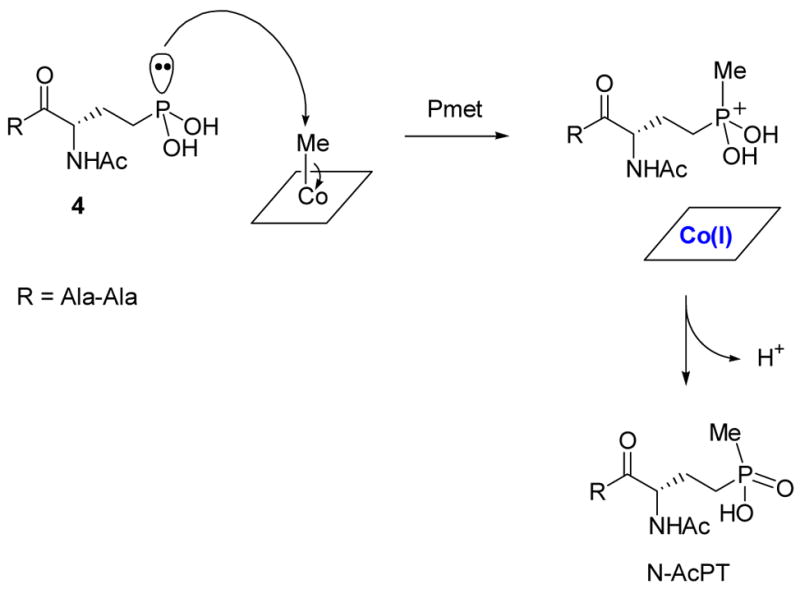
Possible heterolytic mechanism of P-methyltransferase (Pmet).
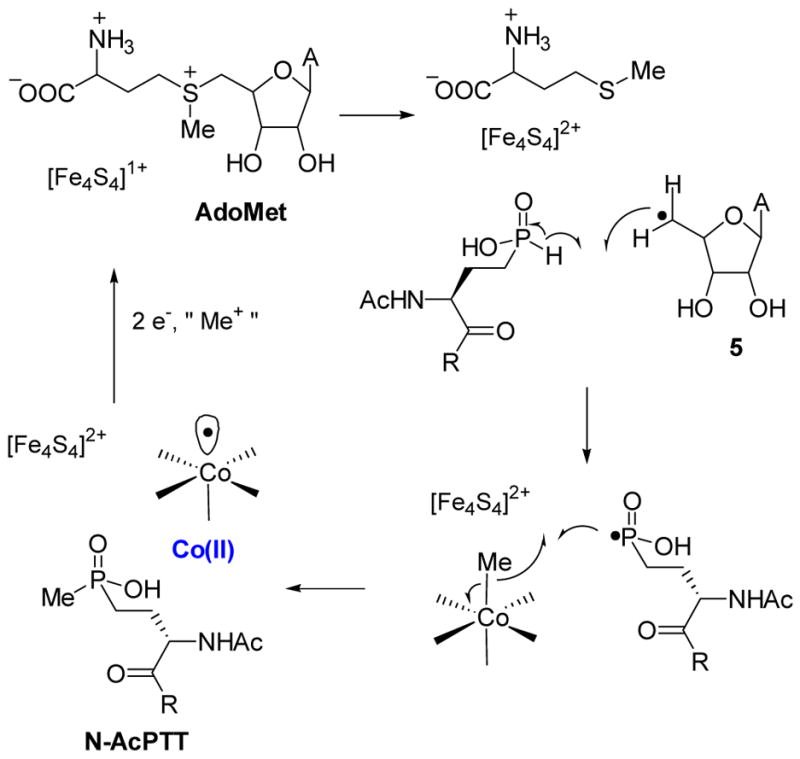
Possible homolytic mechanism of P-methyltransferase (Pmet).
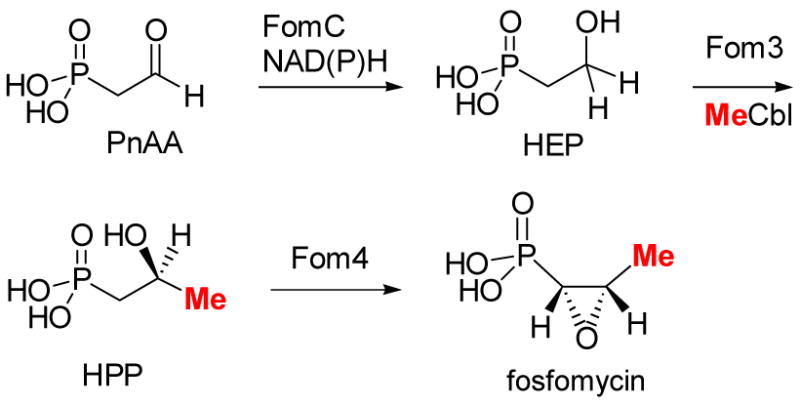
Revised biosynthetic pathway of fosfomycin
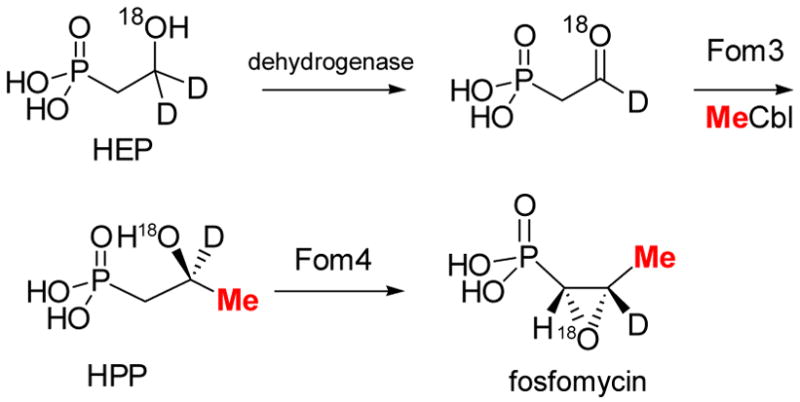
Labeling studies on fosfomycin biosynthesis.
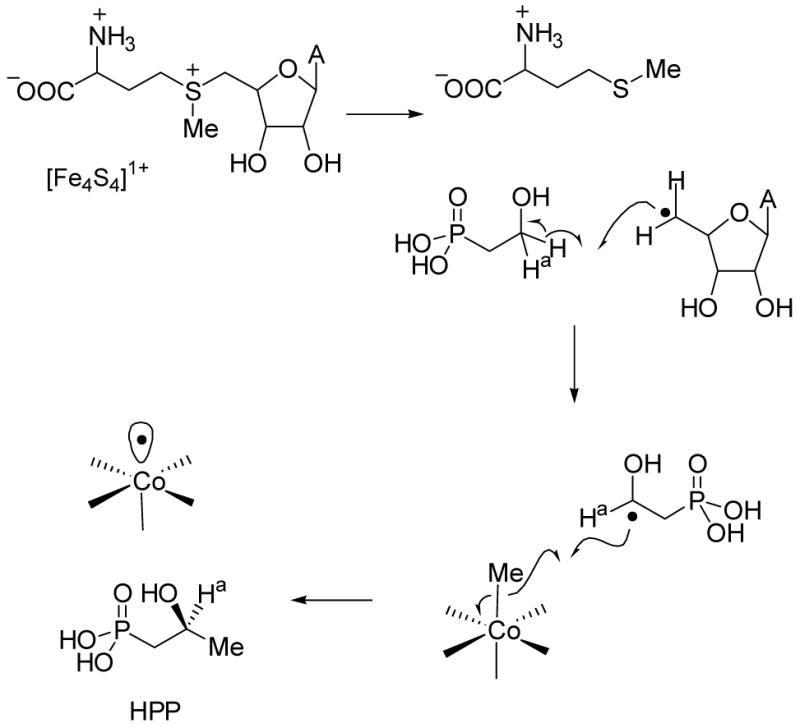
Proposed mechanism for Fom3.
Similar articles
-
First evidence of production of the lantibiotic nisin P.Sci Rep. 2020 Feb 28;10(1):3738. doi: 10.1038/s41598-020-60623-0. Sci Rep. 2020. PMID: 32111904 Free PMC article.
-
Mechanistic dissection of the enzyme complexes involved in biosynthesis of lacticin 3147 and nisin.Appl Environ Microbiol. 2008 Nov;74(21):6591-7. doi: 10.1128/AEM.01334-08. Epub 2008 Sep 12. Appl Environ Microbiol. 2008. PMID: 18791001 Free PMC article.
-
Nisin and lacticin 481 coproduction by Lactococcus lactis strains isolated from raw ewes' milk.J Dairy Sci. 2009 Oct;92(10):4805-11. doi: 10.3168/jds.2009-2237. J Dairy Sci. 2009. PMID: 19762795
-
Bacteriocins: safe, natural antimicrobials for food preservation.Int J Food Microbiol. 2001 Dec 4;71(1):1-20. doi: 10.1016/s0168-1605(01)00560-8. Int J Food Microbiol. 2001. PMID: 11764886 Review.
-
Bioengineering of the model lantibiotic nisin.Bioengineered. 2015;6(4):187-92. doi: 10.1080/21655979.2015.1049781. Epub 2015 May 13. Bioengineered. 2015. PMID: 25970137 Free PMC article. Review.
Cited by
-
Different biosynthetic pathways to fosfomycin in Pseudomonas syringae and Streptomyces species.Antimicrob Agents Chemother. 2012 Aug;56(8):4175-83. doi: 10.1128/AAC.06478-11. Epub 2012 May 21. Antimicrob Agents Chemother. 2012. PMID: 22615277 Free PMC article.
-
Significant increase of oxidase activity through the genetic incorporation of a tyrosine-histidine cross-link in a myoglobin model of heme-copper oxidase.Angew Chem Int Ed Engl. 2012 Apr 27;51(18):4312-6. doi: 10.1002/anie.201108756. Epub 2012 Mar 12. Angew Chem Int Ed Engl. 2012. PMID: 22411709 Free PMC article.
-
Vitamin B12 in the spotlight again.Curr Opin Chem Biol. 2017 Apr;37:63-70. doi: 10.1016/j.cbpa.2017.01.013. Epub 2017 Feb 3. Curr Opin Chem Biol. 2017. PMID: 28167430 Free PMC article. Review.
-
Stereochemical and Mechanistic Investigation of the Reaction Catalyzed by Fom3 from Streptomyces fradiae, a Cobalamin-Dependent Radical S-Adenosylmethionine Methylase.Biochemistry. 2018 Aug 21;57(33):4972-4984. doi: 10.1021/acs.biochem.8b00693. Epub 2018 Aug 9. Biochemistry. 2018. PMID: 30036047 Free PMC article.
-
Radical S-adenosylmethionine enzymes.Chem Rev. 2014 Apr 23;114(8):4229-317. doi: 10.1021/cr4004709. Epub 2014 Jan 29. Chem Rev. 2014. PMID: 24476342 Free PMC article. Review. No abstract available.
References
-
- Li YM, Milne JC, Madison LL, Kolter R, Walsh CT. Science. 1996;274:1188–93. - PubMed
-
- Kuzuyama T, Hidaka T, Kamigiri K, Imai S, Seto H. J Antibiot. 1992;45:1812–4. - PubMed
- Kamigiri K, Hidaka T, Imai S, Murakami T, Seto H. J Antibiot. 1992;45:781–7. - PubMed
- Hidaka T, Goda M, Kuzuyama T, Takei N, Kidaka M, Seto H. Mol Gen Genet. 1995;249:274–280. - PubMed
- Hidaka T, Hidaka M, Kuzuyama T, Seto H. Gene. 1995;158:149–150. - PubMed
- Seto H, Kuzuyama T. Nat Prod Rep. 1999;16:589–596. - PubMed
- Seto H, Hidaka T, Kuzuyama T, Shibahara S, Usui T, Sakanaka O, Imai S. J Antibiot. 1991;44:1286–8. - PubMed
-
- Hammerschmidt F. Angew Chem Int Ed Engl. 1994;33:341–342.
- Hammerschmidt F, Kählig H. J Org Chem. 1991;56:2364–2370.
-
- Banerjee R, editor. The Chemistry and Biochemistry of B12. Wiley; New York: 1999.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
