

Single Holliday junctions are intermediates of meiotic recombination
- PMID: 17174892
- PMCID: PMC2803030
- DOI: 10.1016/j.cell.2006.09.050
Single Holliday junctions are intermediates of meiotic recombination
Abstract
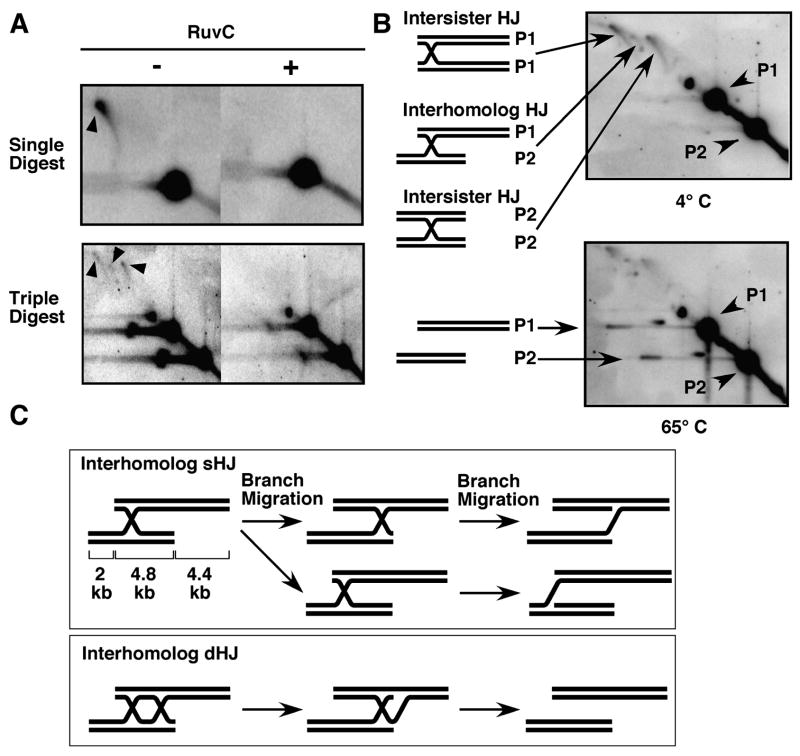
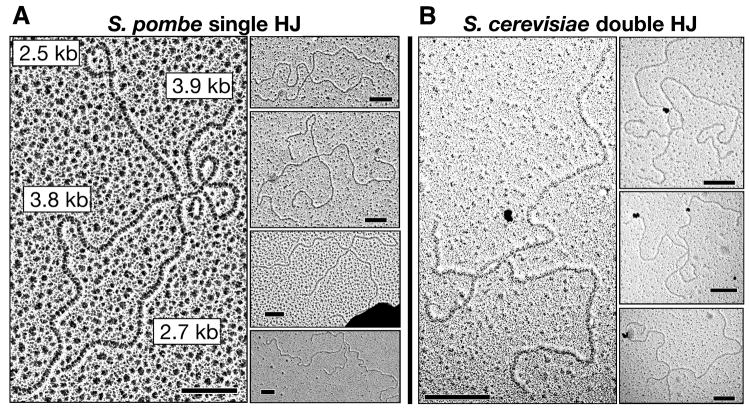
Crossing-over between homologous chromosomes facilitates their accurate segregation at the first division of meiosis. Current models for crossing-over invoke an intermediate in which homologs are connected by two crossed-strand structures called Holliday junctions. Such double Holliday junctions are a prominent intermediate in Saccharomyces cerevisiae meiosis, where they form preferentially between homologs rather than between sister chromatids. In sharp contrast, we find that single Holliday junctions are the predominant intermediate in Schizosaccharomyces pombe meiosis. Furthermore, these single Holliday junctions arise preferentially between sister chromatids rather than between homologs. We show that Mus81 is required for Holliday junction resolution, providing further in vivo evidence that the structure-specific endonuclease Mus81-Eme1 is a Holliday junction resolvase. To reconcile these observations, we present a unifying recombination model applicable for both meiosis and mitosis in which single Holliday junctions arise from single- or double-strand breaks, lesions postulated by previous models to initiate recombination.
Figures




Comment in
-
Multiple mechanisms of meiotic recombination.Cell. 2006 Dec 15;127(6):1095-7. doi: 10.1016/j.cell.2006.11.029. Cell. 2006. PMID: 17174887
References
-
- Allers T, Lichten M. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell. 2001;106:47–57. - PubMed
-
- Bell LR, Byers B. Homologous association of chromosomal DNA during yeast meiosis. Cold Spring Harb Symp Quant Biol. 1983;47:829–840. - PubMed
-
- Benson FE, West SC. Substrate specificity of the Escherichia coli RuvC protein. Resolution of three- and four-stranded recombination intermediates. J Biol Chem. 1994;269:5195–5201. - PubMed
-
- Boddy MN, Gaillard PH, McDonald WH, Shanahan P, Yates JR, 3rd, Russell P. Mus81-Eme1 are essential components of a Holliday junction resolvase. Cell. 2001;107:537–548. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
