

Dynamic role of postsynaptic caspase-3 and BIRC4 in zebra finch song-response habituation
- PMID: 17178408
- PMCID: PMC1847391
- DOI: 10.1016/j.neuron.2006.10.033
Dynamic role of postsynaptic caspase-3 and BIRC4 in zebra finch song-response habituation
Abstract
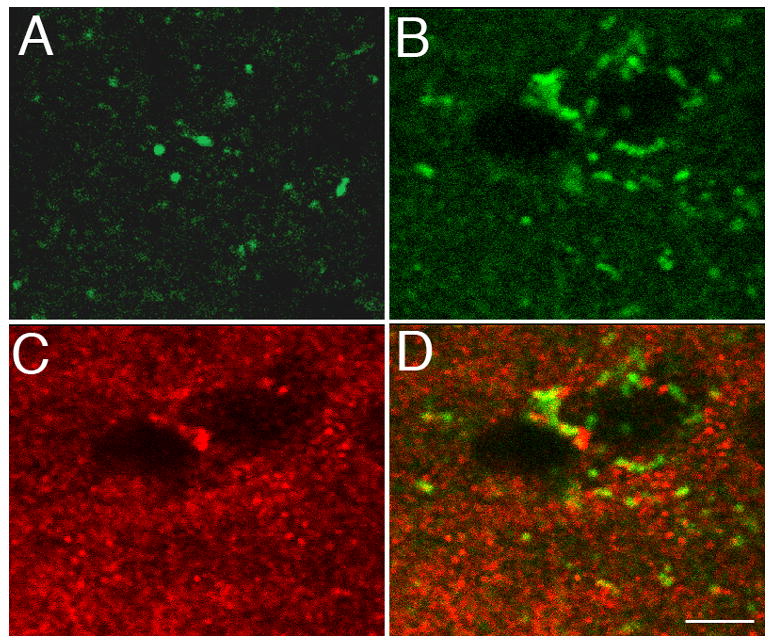
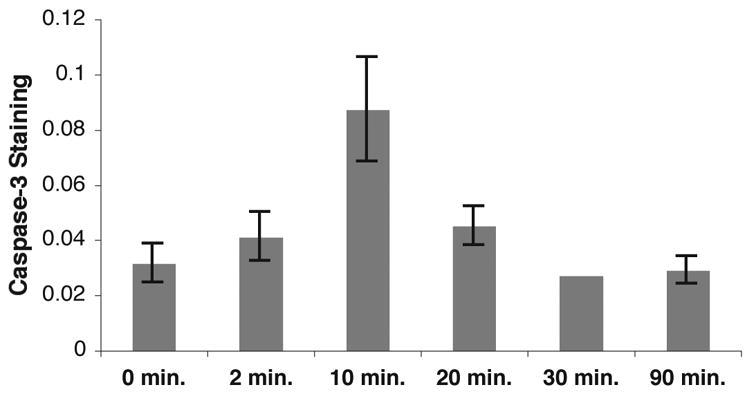
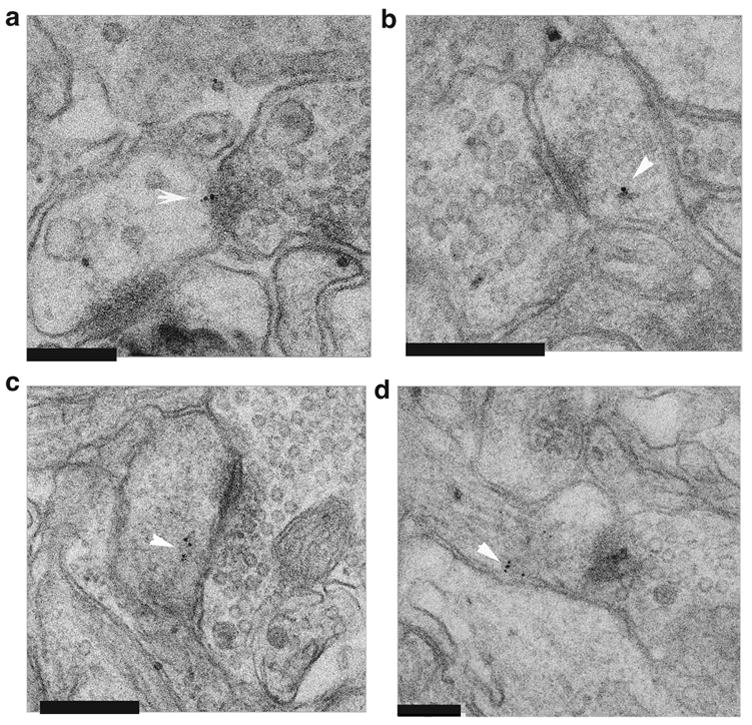
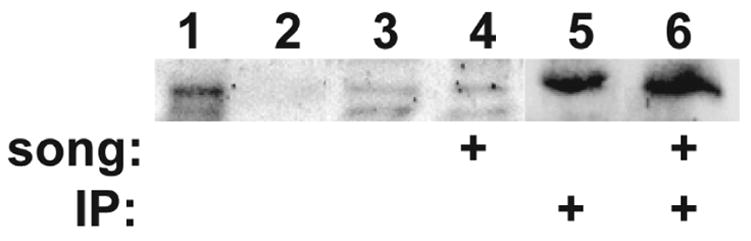
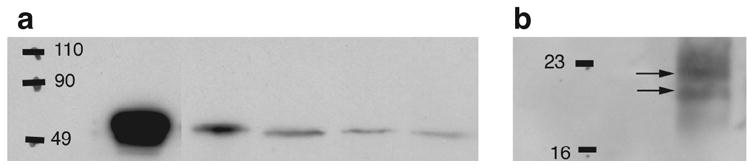
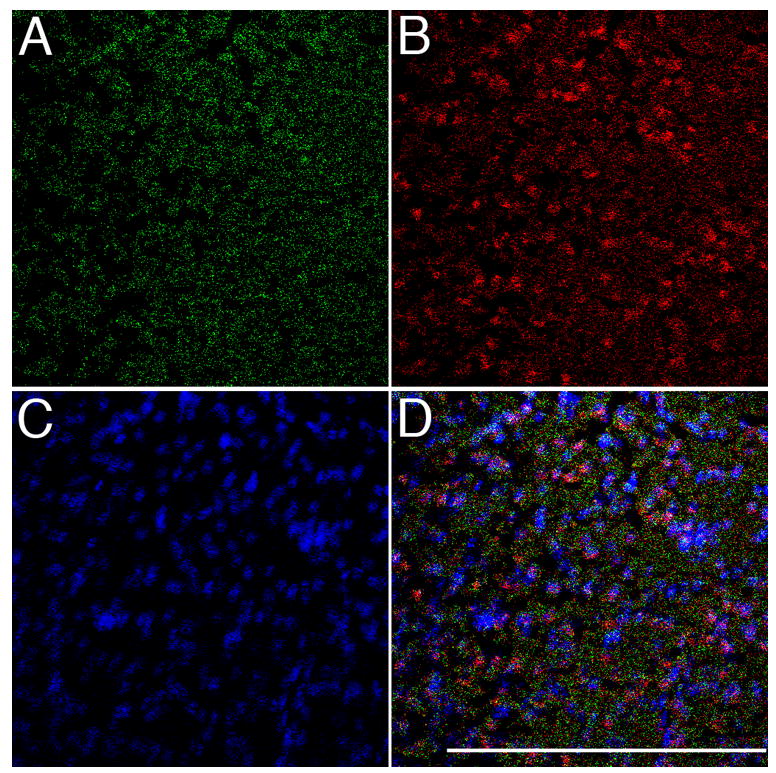
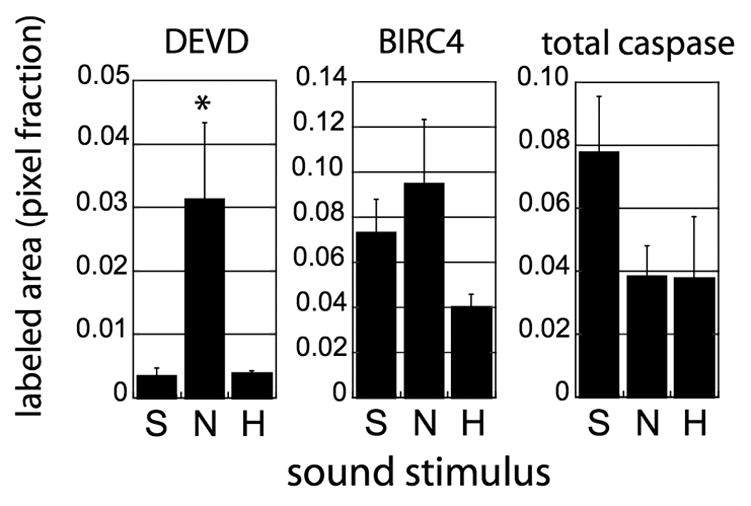
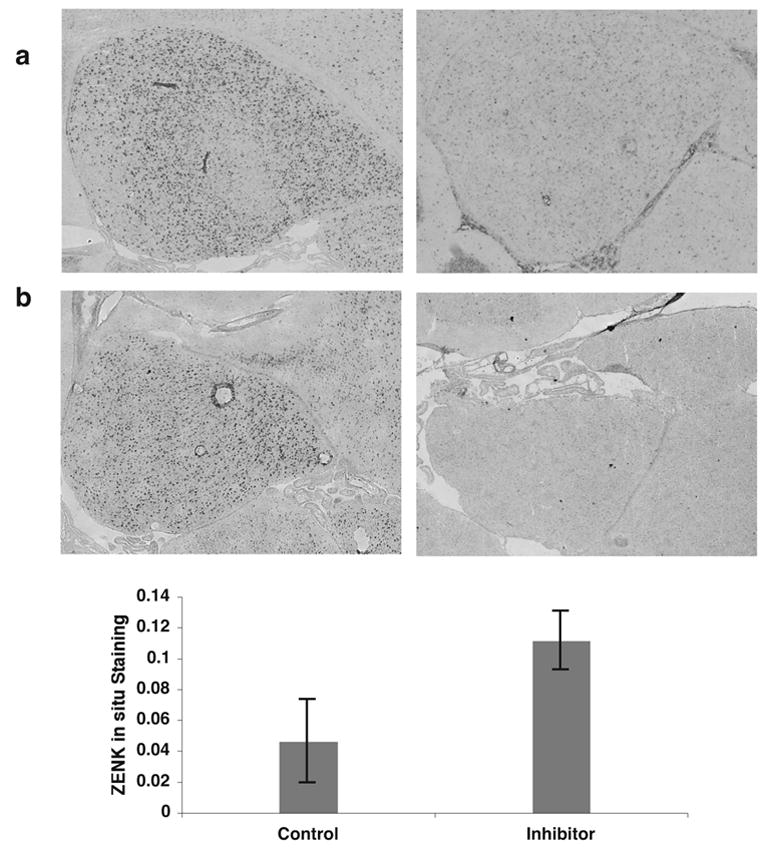
Activation of the protease caspase-3 is commonly thought to cause apoptotic cell death. Here, we show that caspase-3 activity is regulated at postsynaptic sites in brain following stimuli associated with memory (neural activation and subsequent response habituation) instead of cell death. In the zebra finch auditory forebrain, the concentration of caspase-3 active sites increases briefly within minutes after exposure to tape-recorded birdsong. With confocal and immunoelectron microscopy, we localize the activated enzyme to dendritic spines. The activated caspase-3 protein is present even in unstimulated brain but bound to an endogenous inhibitor, BIRC4 (xIAP), suggesting a mechanism for rapid release and sequestering at specific synaptic sites. Caspase-3 activity is necessary to consolidate a persistent physiological trace of the song stimulus, as demonstrated using pharmacological interference and the zenk gene habituation assay. Thus, the brain appears to have adapted a core component of cell death machinery to serve a unique role in learning and memory.
Figures

References
-
- Alvarez-Buylla A, Nottebohm F. Migration of young neurons in adult avian brain. Nature. 1988;335:353–354. - PubMed
-
- Boehning D, Patterson RL, Sedaghat L, Glebova NO, Kurosaki T, Snyder SH. Cytochrome c binds to inositol (1,4,5) trisphosphate receptors, amplifying calcium-dependent apoptosis. Nat Cell Biol. 2003;5:1051–1061. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
