

Coactivating signals for the hepatic lymphocyte gamma interferon response to Francisella tularensis
- PMID: 17178781
- PMCID: PMC1828575
- DOI: 10.1128/IAI.01203-06
Coactivating signals for the hepatic lymphocyte gamma interferon response to Francisella tularensis
Abstract
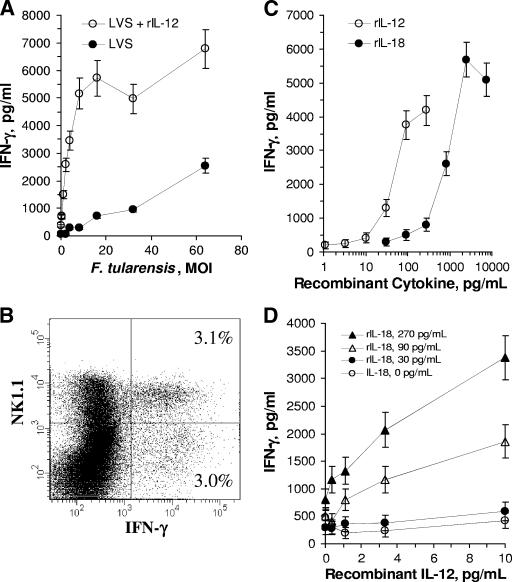
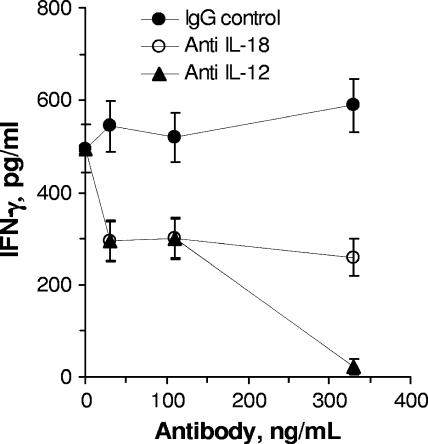
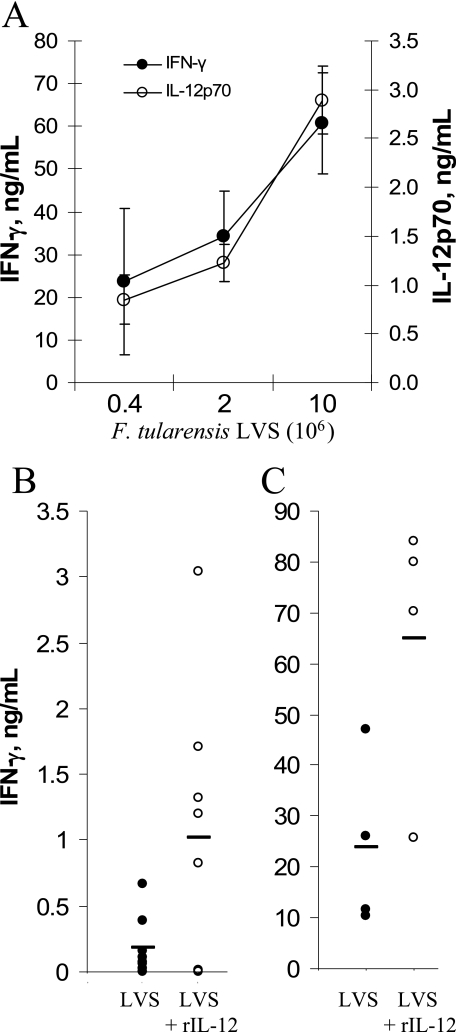
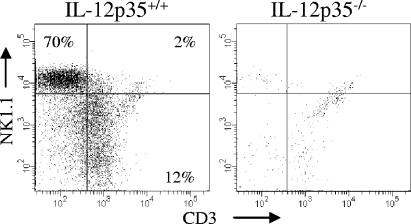
The facultative intracellular bacterium Francisella tularensis is capable of causing systemic infections in various hosts, including mice and humans. The liver is a major secondary site of F. tularensis infection, but hepatic immune responses to the pathogen remain poorly defined. Immune protection against the pathogen is thought to depend on the cytokine gamma interferon (IFN-gamma), but the cellular basis for this response has not been characterized. Here we report that natural killer cells from the livers of naïve uninfected mice produced IFN-gamma when challenged with live bacteria in vitro and that the responses were greatly increased by coactivation of the cells with either recombinant interleukin-12 (IL-12) or IL-18. Moreover, the two cytokines had strong synergistic effects on IFN-gamma induction. Neutralizing antibodies to either IL-12 or IL-18 inhibited IFN-gamma production in vitro, and mice deficient in the p35 subunit of IL-12 failed to show IFN-gamma responses to bacterial challenge either in vitro or in vivo. Clinical isolates of highly virulent type A Francisella tularensis subsp. tularensis organisms were comparable to the live attenuated vaccine strain of Francisella tularensis subsp. holarctica in their ability to induce IL-12 and IFN-gamma expression. These findings demonstrate that cells capable of mounting IFN-gamma responses to F. tularensis are resident within the livers of uninfected mice and depend on coactivation by IL-12 and IL-18 for optimum responses.
Figures
Similar articles
-
Diverse myeloid and lymphoid cell subpopulations produce gamma interferon during early innate immune responses to Francisella tularensis live vaccine strain.Infect Immun. 2008 Sep;76(9):4311-21. doi: 10.1128/IAI.00514-08. Epub 2008 Jun 23. Infect Immun. 2008. PMID: 18573901 Free PMC article.
-
Toll-like receptor 2 controls the gamma interferon response to Francisella tularensis by mouse liver lymphocytes.Infect Immun. 2007 Nov;75(11):5338-45. doi: 10.1128/IAI.00561-07. Epub 2007 Sep 4. Infect Immun. 2007. PMID: 17785474 Free PMC article.
-
Lung CD4-CD8- double-negative T cells are prominent producers of IL-17A and IFN-gamma during primary respiratory murine infection with Francisella tularensis live vaccine strain.J Immunol. 2010 May 15;184(10):5791-801. doi: 10.4049/jimmunol.1000362. Epub 2010 Apr 14. J Immunol. 2010. PMID: 20393138
-
Francisella tularensis: activation of the inflammasome.Ann N Y Acad Sci. 2007 Jun;1105:219-37. doi: 10.1196/annals.1409.005. Epub 2007 Mar 29. Ann N Y Acad Sci. 2007. PMID: 17395724 Review.
-
Francisella and Antibodies.Microorganisms. 2021 Oct 12;9(10):2136. doi: 10.3390/microorganisms9102136. Microorganisms. 2021. PMID: 34683457 Free PMC article. Review.
Cited by
-
Working toward the future: insights into Francisella tularensis pathogenesis and vaccine development.Microbiol Mol Biol Rev. 2009 Dec;73(4):684-711. doi: 10.1128/MMBR.00028-09. Microbiol Mol Biol Rev. 2009. PMID: 19946137 Free PMC article. Review.
-
Diverse myeloid and lymphoid cell subpopulations produce gamma interferon during early innate immune responses to Francisella tularensis live vaccine strain.Infect Immun. 2008 Sep;76(9):4311-21. doi: 10.1128/IAI.00514-08. Epub 2008 Jun 23. Infect Immun. 2008. PMID: 18573901 Free PMC article.
-
Cytokine response in Balb/c mice infected with Francisella tularensis LVS and the Pohang isolate.J Vet Sci. 2008 Sep;9(3):309-15. doi: 10.4142/jvs.2008.9.3.309. J Vet Sci. 2008. PMID: 18716452 Free PMC article.
-
Francisella tularensis induces extensive caspase-3 activation and apoptotic cell death in the tissues of infected mice.Infect Immun. 2009 Nov;77(11):4827-36. doi: 10.1128/IAI.00246-09. Epub 2009 Aug 24. Infect Immun. 2009. PMID: 19703976 Free PMC article.
-
Phenotypic, morphological, and functional heterogeneity of splenic immature myeloid cells in the host response to tularemia.Infect Immun. 2012 Jul;80(7):2371-81. doi: 10.1128/IAI.00365-12. Epub 2012 Apr 23. Infect Immun. 2012. PMID: 22526678 Free PMC article.
References
-
- Anthony, L. S., E. Ghadirian, F. P. Nestel, and P. A. Kongshavn. 1989. The requirement for gamma interferon in resistance of mice to experimental tularemia. Microb. Pathog. 7:421-428. - PubMed
-
- Anthony, L. S., P. J. Morrissey, and F. E. Nano. 1992. Growth inhibition of Francisella tularensis live vaccine strain by IFN-γ-activated macrophages is mediated by reactive nitrogen intermediates derived from l-arginine metabolism. J. Immunol. 148:1829-1834. - PubMed
-
- Biet, F., C. Locht, and L. Kremer. 2002. Immunoregulatory functions of interleukin 18 and its role in defense against bacterial pathogens. J. Mol. Med. 80:147-162. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
