

Deterministic and stochastic regimes of asexual evolution on rugged fitness landscapes
- PMID: 17179085
- PMCID: PMC1840068
- DOI: 10.1534/genetics.106.067165
Deterministic and stochastic regimes of asexual evolution on rugged fitness landscapes
Abstract
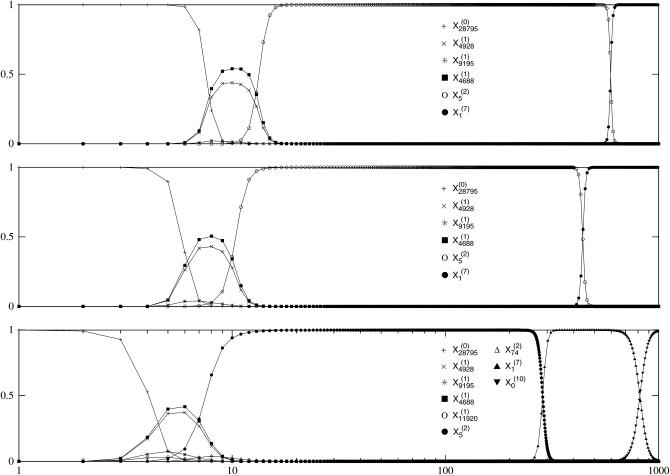
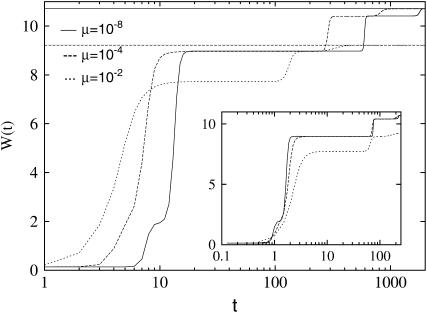
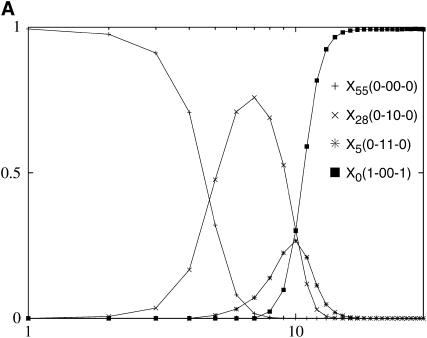
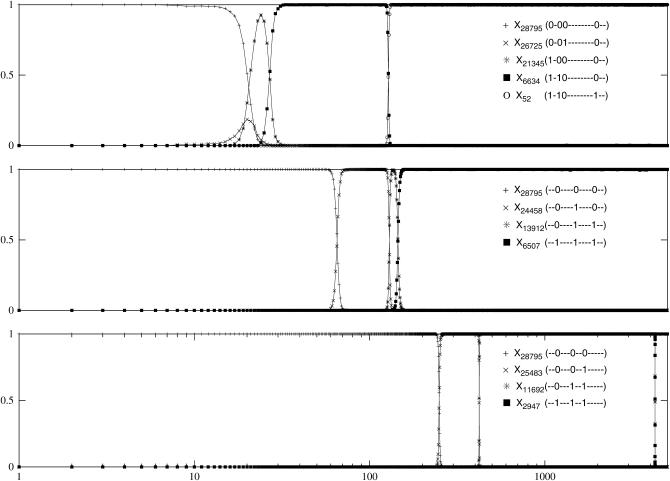
We study the adaptation dynamics of an initially maladapted asexual population with genotypes represented by binary sequences of length L. The population evolves in a maximally rugged fitness landscape with a large number of local optima. We find that whether the evolutionary trajectory is deterministic or stochastic depends on the effective mutational distance d(eff) up to which the population can spread in genotype space. For d(eff) = L, the deterministic quasi-species theory operates while for d(eff) < 1, the evolution is completely stochastic. Between these two limiting cases, the dynamics are described by a local quasi-species theory below a crossover time T(x) while above T(x) the population gets trapped at a local fitness peak and manages to find a better peak via either stochastic tunneling or double mutations. In the stochastic regime d(eff) < 1, we identify two subregimes associated with clonal interference and uphill adaptive walks, respectively. We argue that our findings are relevant to the interpretation of evolution experiments with microbial populations.
Figures



Similar articles
-
The impact of population size on the evolution of asexual microbes on smooth versus rugged fitness landscapes.BMC Evol Biol. 2009 Sep 18;9:236. doi: 10.1186/1471-2148-9-236. BMC Evol Biol. 2009. PMID: 19765292 Free PMC article.
-
Evolutionary dynamics of the most populated genotype on rugged fitness landscapes.Phys Rev E Stat Nonlin Soft Matter Phys. 2007 Sep;76(3 Pt 1):031922. doi: 10.1103/PhysRevE.76.031922. Epub 2007 Sep 26. Phys Rev E Stat Nonlin Soft Matter Phys. 2007. PMID: 17930286
-
The effect of bacterial recombination on adaptation on fitness landscapes with limited peak accessibility.PLoS Comput Biol. 2012;8(10):e1002735. doi: 10.1371/journal.pcbi.1002735. Epub 2012 Oct 25. PLoS Comput Biol. 2012. PMID: 23133344 Free PMC article.
-
Beneficial mutations and the dynamics of adaptation in asexual populations.Philos Trans R Soc Lond B Biol Sci. 2010 Apr 27;365(1544):1255-63. doi: 10.1098/rstb.2009.0290. Philos Trans R Soc Lond B Biol Sci. 2010. PMID: 20308101 Free PMC article. Review.
-
Evolutionary genetics: a piggyback ride to adaptation and diversity.Curr Biol. 2007 May 1;17(9):R333-5. doi: 10.1016/j.cub.2007.02.042. Curr Biol. 2007. PMID: 17470349 Review.
Cited by
-
Predictability of evolution depends nonmonotonically on population size.Proc Natl Acad Sci U S A. 2013 Jan 8;110(2):571-6. doi: 10.1073/pnas.1213613110. Epub 2012 Dec 24. Proc Natl Acad Sci U S A. 2013. PMID: 23267075 Free PMC article.
-
Evolutionary graph theory beyond single mutation dynamics: on how network-structured populations cross fitness landscapes.Genetics. 2024 Jun 5;227(2):iyae055. doi: 10.1093/genetics/iyae055. Genetics. 2024. PMID: 38639307 Free PMC article.
-
Efficient escape from local optima in a highly rugged fitness landscape by evolving RNA virus populations.Proc Biol Sci. 2016 Aug 17;283(1836):20160984. doi: 10.1098/rspb.2016.0984. Proc Biol Sci. 2016. PMID: 27534955 Free PMC article.
-
Inference for one-step beneficial mutations using next generation sequencing.Stat Appl Genet Mol Biol. 2015 Feb;14(1):65-81. doi: 10.1515/sagmb-2014-0030. Stat Appl Genet Mol Biol. 2015. PMID: 25720101 Free PMC article.
-
The impact of population size on the evolution of asexual microbes on smooth versus rugged fitness landscapes.BMC Evol Biol. 2009 Sep 18;9:236. doi: 10.1186/1471-2148-9-236. BMC Evol Biol. 2009. PMID: 19765292 Free PMC article.
References
-
- Baake, E., and W. Gabriel, 2000. Biological evolution through mutation, selection, and drift: an introductory review, pp. 203–264 in Annual Reviews of Computational Physics VII, edited by D. Stauffer. World Scientific, Singapore.
-
- Burch, C. L., and L. Chao, 2000. Evolvability of an RNA virus is determined by its mutational neighbourhood. Nature 406: 625–628. - PubMed
-
- David, H. A., 1970. Order Statistics. Wiley, New York.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
