

Gene function and expression level influence the insertion/fixation dynamics of distinct transposon families in mammalian introns
- PMID: 17181857
- PMCID: PMC1794433
- DOI: 10.1186/gb-2006-7-12-r120
Gene function and expression level influence the insertion/fixation dynamics of distinct transposon families in mammalian introns
Abstract
Background: Transposable elements (TEs) represent more than 45% of the human and mouse genomes. Both parasitic and mutualistic features have been shown to apply to the host-TE relationship but a comprehensive scenario of the forces driving TE fixation within mammalian genes is still missing.
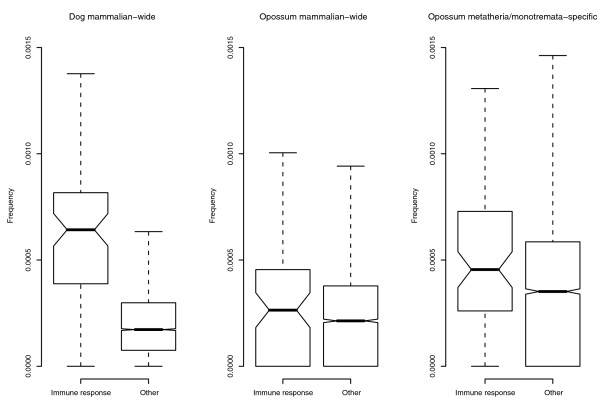
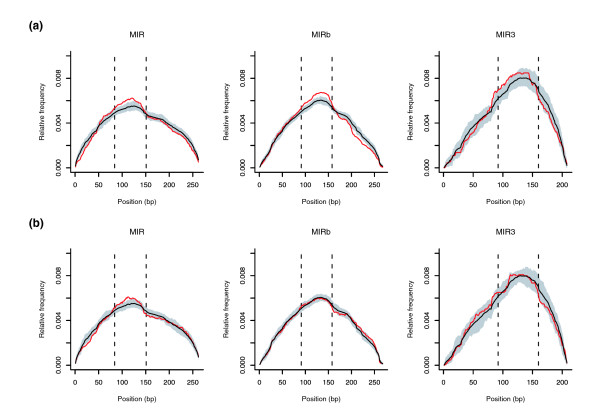
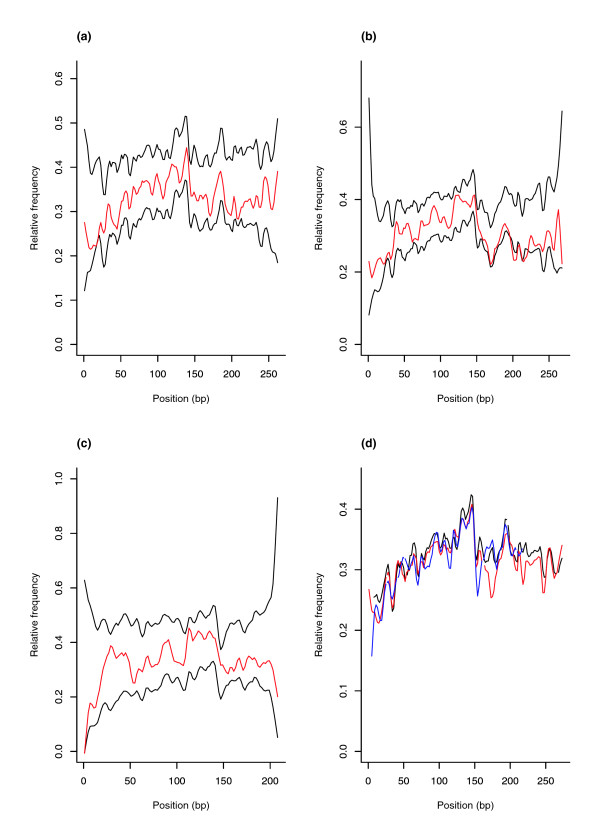
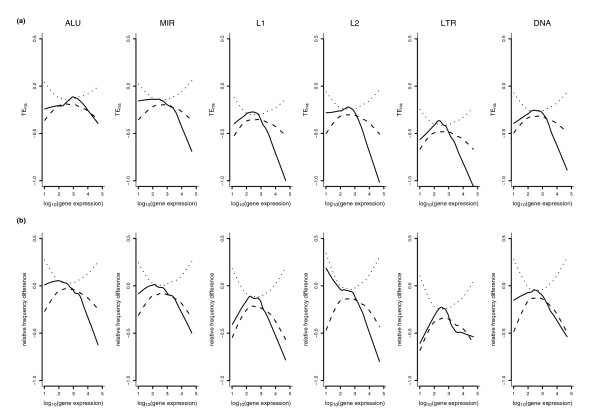
Results: We show that intronic multispecies conserved sequences (MCSs) have been affecting TE integration frequency over time. We verify that a selective economizing pressure has been acting on TEs to decrease their frequency in highly expressed genes. After correcting for GC content, MCS density and intron size, we identified TE-enriched and TE-depleted gene categories. In addition to developmental regulators and transcription factors, TE-depleted regions encompass loci that might require subtle regulation of transcript levels or precise activation timing, such as growth factors, cytokines, hormones, and genes involved in the immune response. The latter, despite having reduced frequencies of most TE types, are significantly enriched in mammalian-wide interspersed repeats (MIRs). Analysis of orthologous genes indicated that MIR over-representation also occurs in dog and opossum immune response genes, suggesting, given the partially independent origin of MIR sequences in eutheria and metatheria, the evolutionary conservation of a specific function for MIRs located in these loci. Consistently, the core MIR sequence is over-represented in defense response genes compared to the background intronic frequency.
Conclusion: Our data indicate that gene function, expression level, and sequence conservation influence TE insertion/fixation in mammalian introns. Moreover, we provide the first report showing that a specific TE family is evolutionarily associated with a gene function category.
Figures
Similar articles
-
Fixation of conserved sequences shapes human intron size and influences transposon-insertion dynamics.Trends Genet. 2005 Sep;21(9):484-8. doi: 10.1016/j.tig.2005.06.009. Trends Genet. 2005. PMID: 16005101
-
Distributions of transposable elements reveal hazardous zones in mammalian introns.PLoS Comput Biol. 2011 May;7(5):e1002046. doi: 10.1371/journal.pcbi.1002046. Epub 2011 May 5. PLoS Comput Biol. 2011. PMID: 21573203 Free PMC article.
-
Analysis of intronic conserved elements indicates that functional complexity might represent a major source of negative selection on non-coding sequences.Hum Mol Genet. 2005 Sep 1;14(17):2533-46. doi: 10.1093/hmg/ddi257. Epub 2005 Jul 21. Hum Mol Genet. 2005. PMID: 16037065
-
Transposable element influences on gene expression in plants.Biochim Biophys Acta Gene Regul Mech. 2017 Jan;1860(1):157-165. doi: 10.1016/j.bbagrm.2016.05.010. Epub 2016 May 25. Biochim Biophys Acta Gene Regul Mech. 2017. PMID: 27235540 Review.
-
TFs for TEs: the transcription factor repertoire of mammalian transposable elements.Genes Dev. 2021 Jan 1;35(1-2):22-39. doi: 10.1101/gad.344473.120. Genes Dev. 2021. PMID: 33397727 Free PMC article. Review.
Cited by
-
Characteristics of transposable element exonization within human and mouse.PLoS One. 2010 Jun 1;5(6):e10907. doi: 10.1371/journal.pone.0010907. PLoS One. 2010. PMID: 20532223 Free PMC article.
-
The ant genomes have been invaded by several types of mariner transposable elements.Naturwissenschaften. 2012 Dec;99(12):1007-20. doi: 10.1007/s00114-012-0982-5. Epub 2012 Oct 25. Naturwissenschaften. 2012. PMID: 23097152
-
The (r)evolution of SINE versus LINE distributions in primate genomes: sex chromosomes are important.Genome Res. 2010 May;20(5):600-13. doi: 10.1101/gr.099044.109. Epub 2010 Mar 10. Genome Res. 2010. PMID: 20219940 Free PMC article.
-
Evolutionary rate of human tissue-specific genes are related with transposable element insertions.Genetica. 2012 Dec;140(10-12):513-23. doi: 10.1007/s10709-013-9700-2. Epub 2013 Jan 22. Genetica. 2012. PMID: 23337972
-
Genomic landscape of human, bat, and ex vivo DNA transposon integrations.Mol Biol Evol. 2014 Jul;31(7):1816-32. doi: 10.1093/molbev/msu138. Epub 2014 Apr 22. Mol Biol Evol. 2014. PMID: 24809961 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
