

The 1',4'-iminopyrimidine tautomer of thiamin diphosphate is poised for catalysis in asymmetric active centers on enzymes
- PMID: 17182735
- PMCID: PMC1765481
- DOI: 10.1073/pnas.0609973104
The 1',4'-iminopyrimidine tautomer of thiamin diphosphate is poised for catalysis in asymmetric active centers on enzymes
Abstract
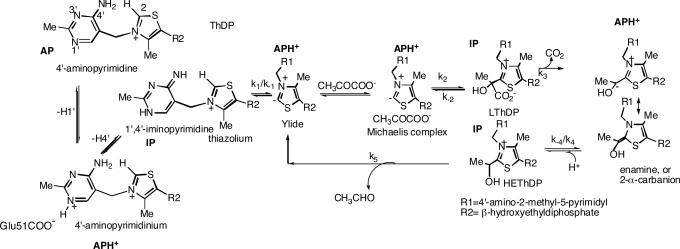
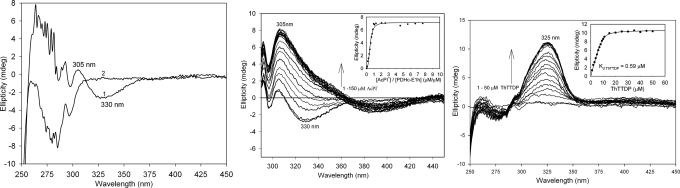
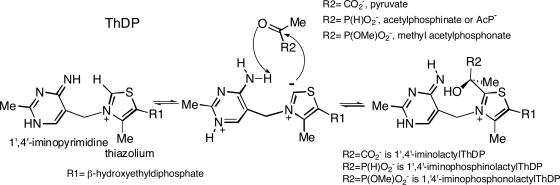
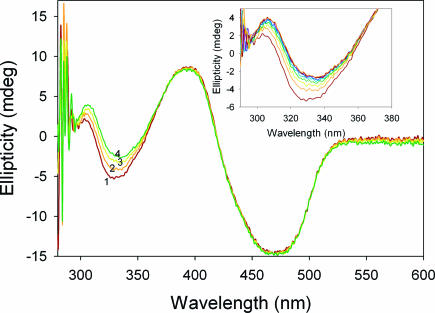
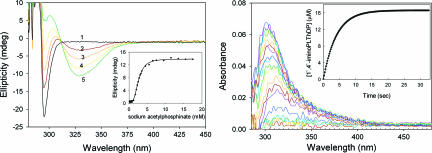
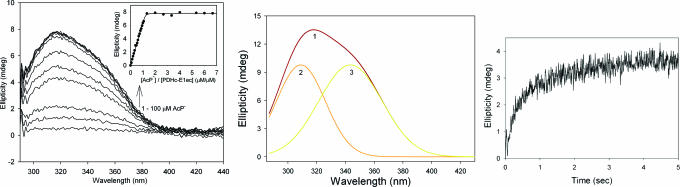
Thiamin diphosphate, a key coenzyme in sugar metabolism, is comprised of the thiazolium and 4'-aminopyrimidine aromatic rings, but only recently has participation of the 4'-aminopyrimidine moiety in catalysis gained wider acceptance. We report the use of electronic spectroscopy to identify the various tautomeric forms of the 4'-aminopyrimidine ring on four thiamin diphosphate enzymes, all of which decarboxylate pyruvate: the E1 component of human pyruvate dehydrogenase complex, the E1 subunit of Escherichia coli pyruvate dehydrogenase complex, yeast pyruvate decarboxylase, and pyruvate oxidase from Lactobacillus plantarum. It is shown that, according to circular dichroism spectroscopy, both the 1',4'-iminopyrimidine and the 4'-aminopyrimidine tautomers coexist on the E1 component of human pyruvate dehydrogenase complex and pyruvate oxidase. Because both tautomers are seen simultaneously, these two enzymes provide excellent evidence for nonidentical active centers (asymmetry) in solution in these multimeric enzymes. Asymmetry of active centers can also be induced upon addition of acetylphosphinate, an excellent electrostatic pyruvate mimic, which participates in an enzyme-catalyzed addition to form a stable adduct, resembling the common predecarboxylation thiamin-bound intermediate, which exists in its 1',4'-iminopyrimidine form. The identification of the 1',4'-iminopyrimidine tautomer on four enzymes is almost certainly applicable to all thiamin diphosphate enzymes: this tautomer is the intramolecular trigger to generate the reactive ylide/carbene at the thiazolium C2 position in the first fundamental step of thiamin catalysis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
