

Gain-of-function mutations in complement factor B are associated with atypical hemolytic uremic syndrome
- PMID: 17182750
- PMCID: PMC1765442
- DOI: 10.1073/pnas.0603420103
Gain-of-function mutations in complement factor B are associated with atypical hemolytic uremic syndrome
Erratum in
- Proc Natl Acad Sci U S A. 2007 Jun 19;104(25):10749
Abstract
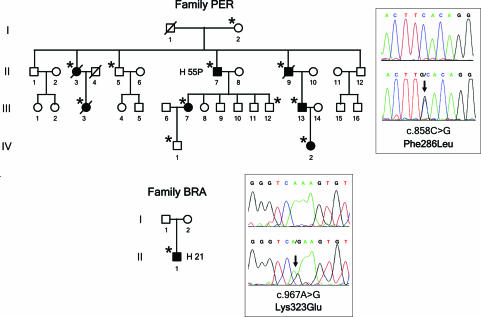
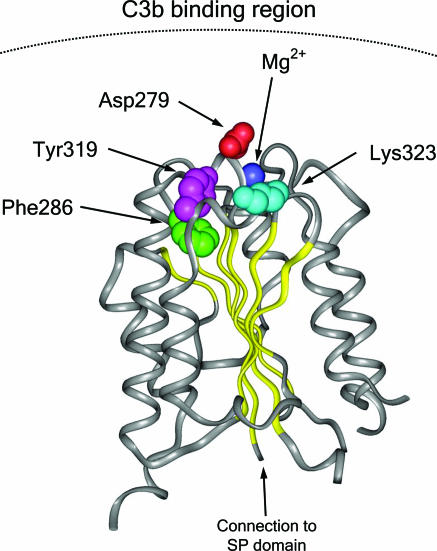
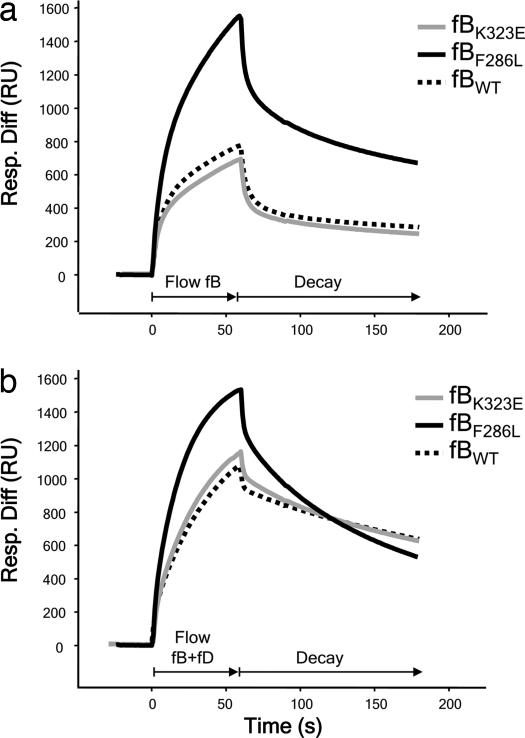
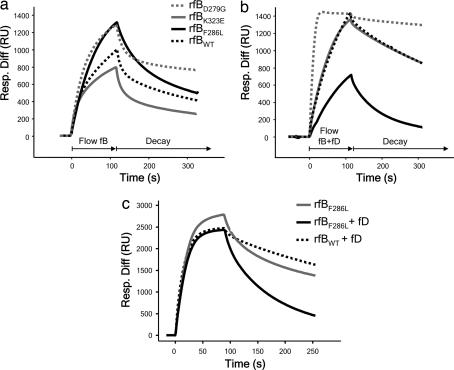
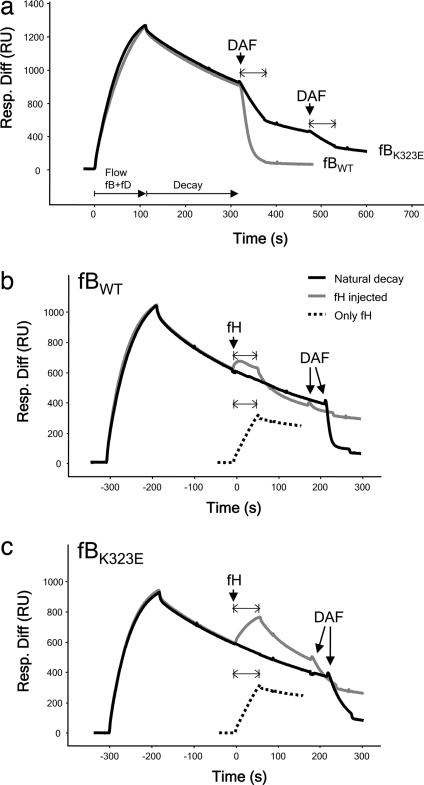
Hemolytic uremic syndrome (HUS) is an important cause of acute renal failure in children. Mutations in one or more genes encoding complement-regulatory proteins have been reported in approximately one-third of nondiarrheal, atypical HUS (aHUS) patients, suggesting a defect in the protection of cell surfaces against complement activation in susceptible individuals. Here, we identified a subgroup of aHUS patients showing persistent activation of the complement alternative pathway and found within this subgroup two families with mutations in the gene encoding factor B (BF), a zymogen that carries the catalytic site of the complement alternative pathway convertase (C3bBb). Functional analyses demonstrated that F286L and K323E aHUS-associated BF mutations are gain-of-function mutations that result in enhanced formation of the C3bBb convertase or increased resistance to inactivation by complement regulators. These data expand our understanding of the genetic factors conferring predisposition to aHUS, demonstrate the critical role of the alternative complement pathway in the pathogenesis of aHUS, and provide support for the use of complement-inhibition therapies to prevent or reduce tissue damage caused by dysregulated complement activation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
