

Local edge detectors: a substrate for fine spatial vision at low temporal frequencies in rabbit retina
- PMID: 17182775
- PMCID: PMC6675005
- DOI: 10.1523/JNEUROSCI.1991-06.2006
Local edge detectors: a substrate for fine spatial vision at low temporal frequencies in rabbit retina
Abstract
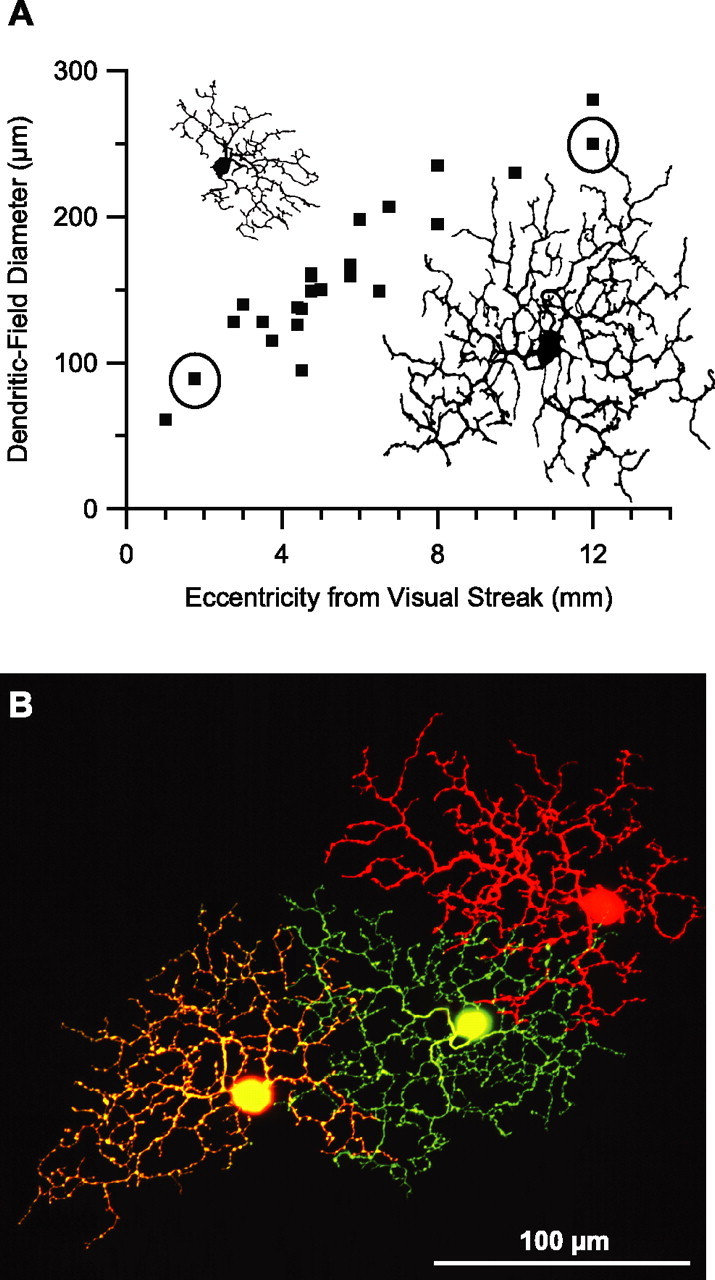
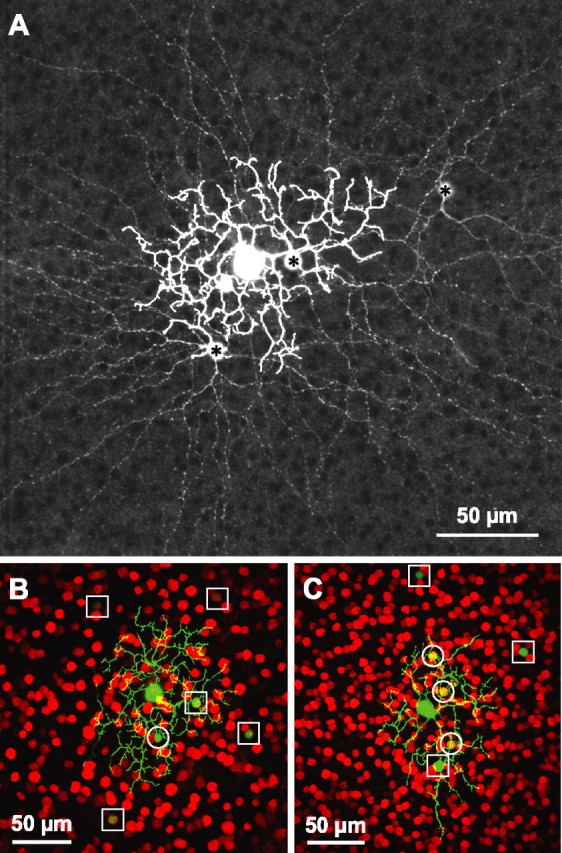
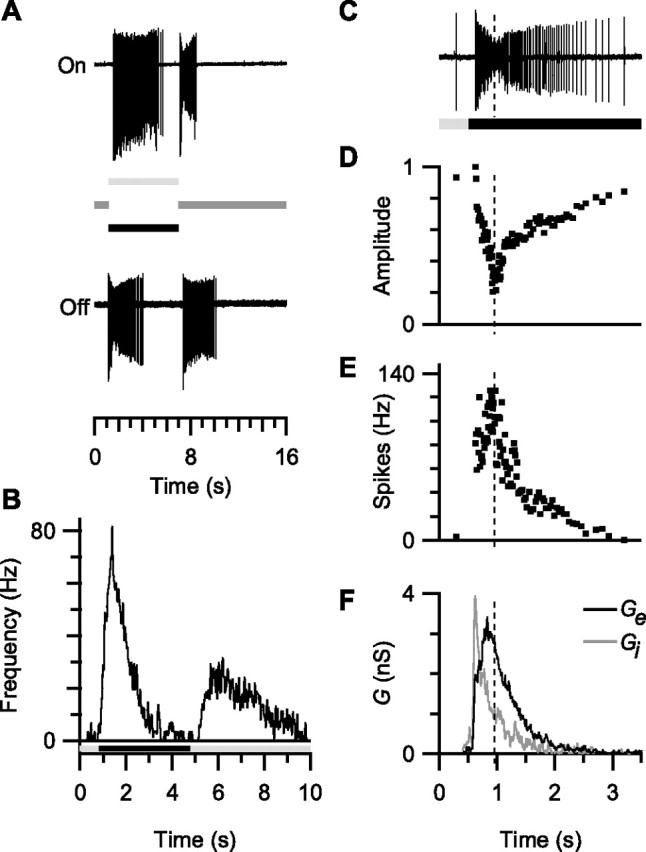
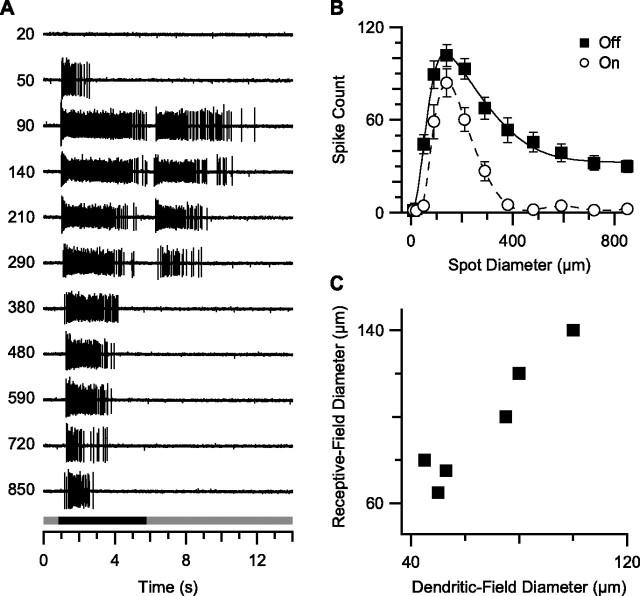
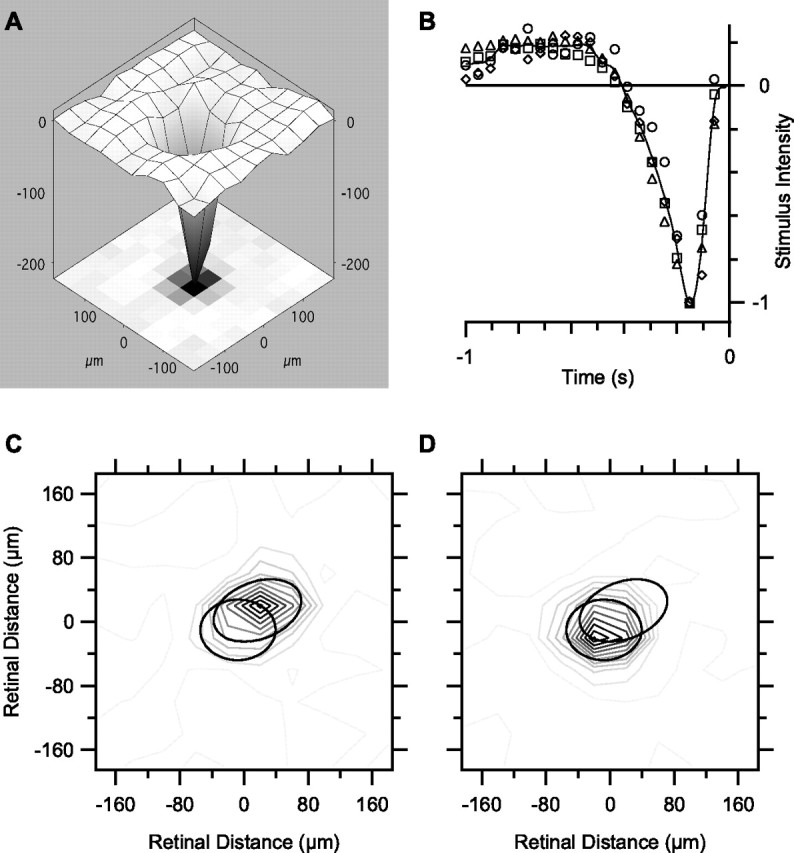
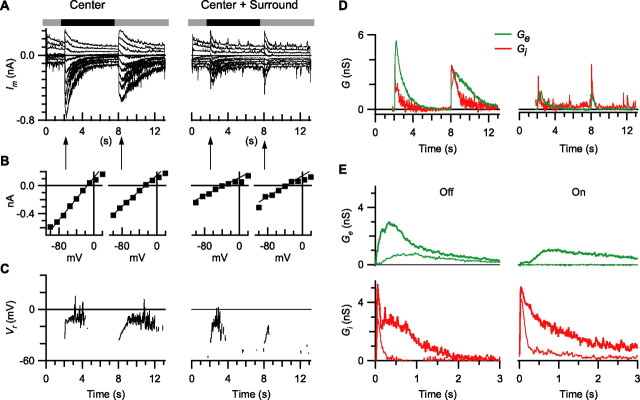
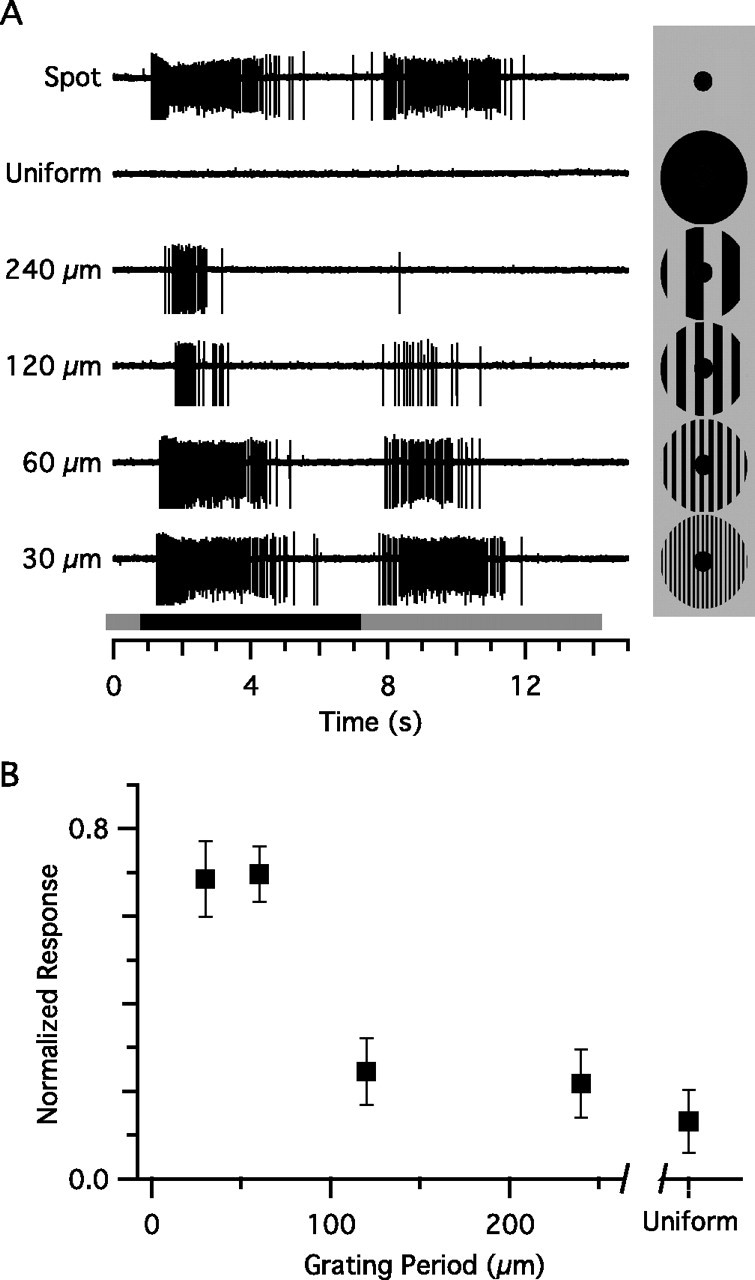
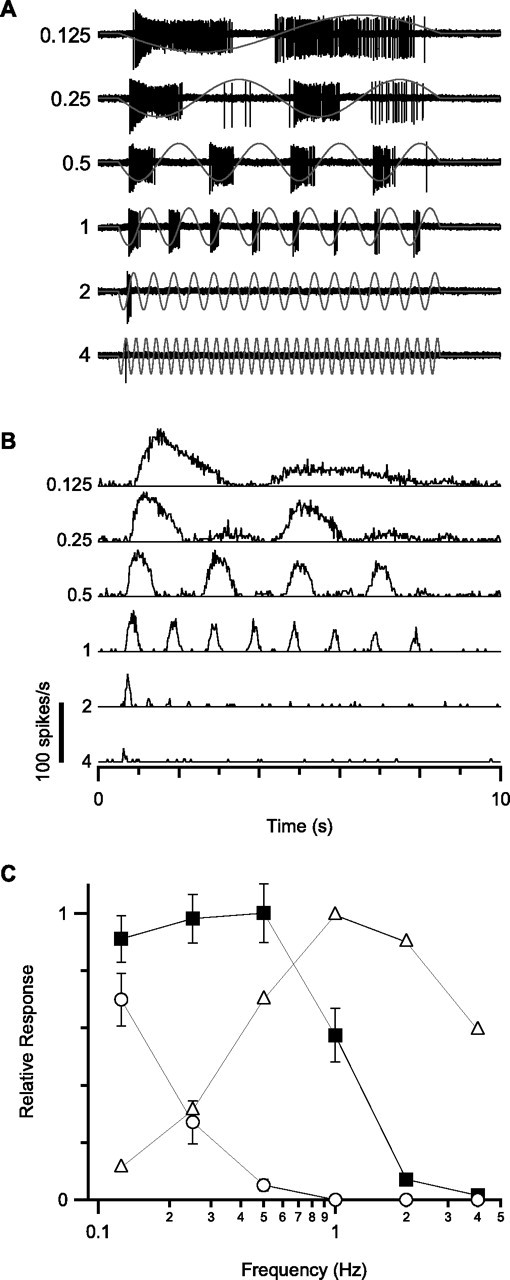
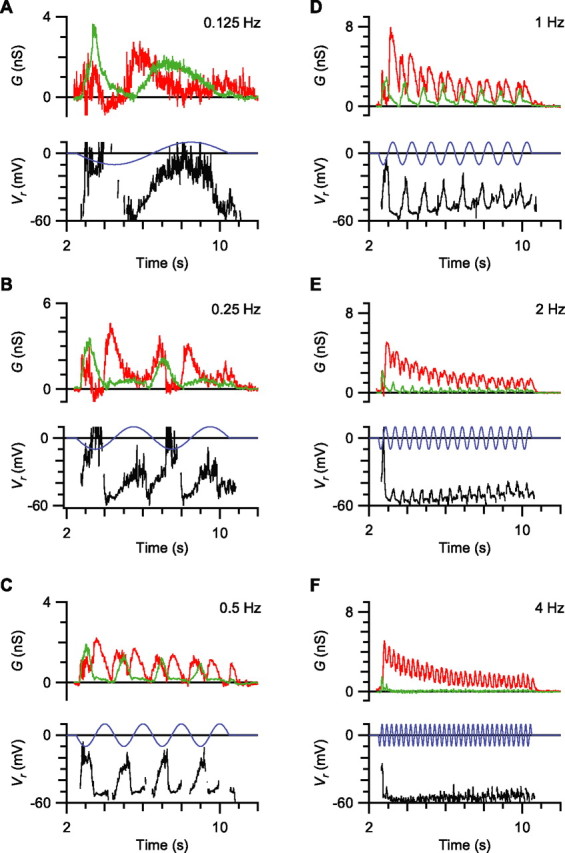
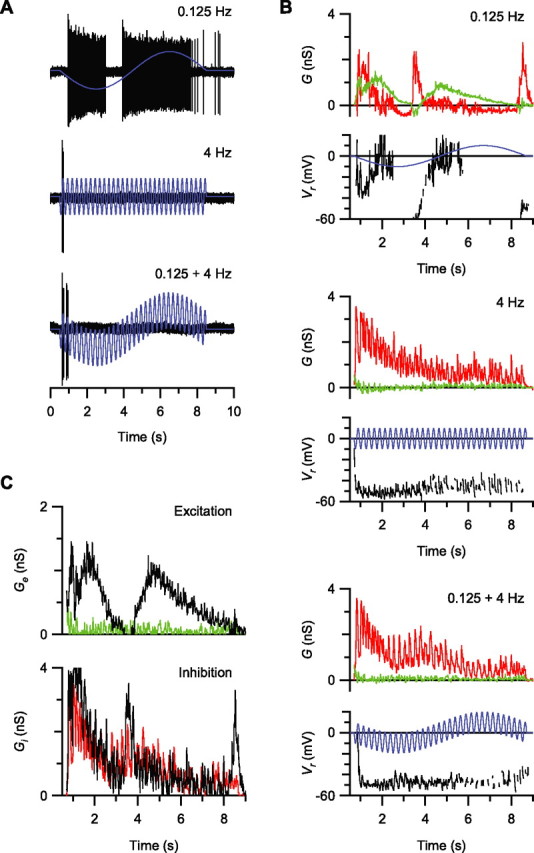
Visual acuity is limited by the size and density of the smallest retinal ganglion cells, which correspond to the midget ganglion cells in primate retina and the beta-ganglion cells in cat retina, both of which have concentric receptive fields that respond at either light-On or light-Off. In contrast, the smallest ganglion cells in the rabbit retina are the local edge detectors (LEDs), which respond to spot illumination at both light-On and light-Off. However, the LEDs do not predominate in the rabbit retina and the question arises, what role do they play in fine spatial vision? We studied the morphology and physiology of LEDs in the isolated rabbit retina and examined how their response properties are shaped by the excitatory and inhibitory inputs. Although the LEDs comprise only approximately 15% of the ganglion cells, neighboring LEDs are separated by 30-40 microm on the visual streak, which is sufficient to account for the grating acuity of the rabbit. The spatial and temporal receptive-field properties of LEDs are generated by distinct inhibitory mechanisms. The strong inhibitory surround acts presynaptically to suppress both the excitation and the inhibition elicited by center stimulation. The temporal properties, characterized by sluggish onset, sustained firing, and low bandwidth, are mediated by the temporal properties of the bipolar cells and by postsynaptic interactions between the excitatory and inhibitory inputs. We propose that the LEDs signal fine spatial detail during visual fixation, when high temporal frequencies are minimal.
Figures
Similar articles
-
Distinct roles for inhibition in spatial and temporal tuning of local edge detectors in the rabbit retina.PLoS One. 2014 Feb 21;9(2):e88560. doi: 10.1371/journal.pone.0088560. eCollection 2014. PLoS One. 2014. PMID: 24586343 Free PMC article.
-
The Synaptic and Morphological Basis of Orientation Selectivity in a Polyaxonal Amacrine Cell of the Rabbit Retina.J Neurosci. 2015 Sep 30;35(39):13336-50. doi: 10.1523/JNEUROSCI.1712-15.2015. J Neurosci. 2015. PMID: 26424882 Free PMC article.
-
Synaptic Mechanisms Generating Orientation Selectivity in the ON Pathway of the Rabbit Retina.J Neurosci. 2016 Mar 16;36(11):3336-49. doi: 10.1523/JNEUROSCI.1432-15.2016. J Neurosci. 2016. PMID: 26985041 Free PMC article.
-
Retinal ganglion cells--spatial organization of the receptive field reduces temporal redundancy.Eur J Neurosci. 2008 Sep;28(5):914-23. doi: 10.1111/j.1460-9568.2008.06394.x. Epub 2008 Aug 8. Eur J Neurosci. 2008. PMID: 18691326 Free PMC article.
-
Diverse inhibitory and excitatory mechanisms shape temporal tuning in transient OFF α ganglion cells in the rabbit retina.J Physiol. 2018 Feb 1;596(3):477-495. doi: 10.1113/JP275195. Epub 2018 Jan 15. J Physiol. 2018. PMID: 29222817 Free PMC article.
Cited by
-
Broad thorny ganglion cells: a candidate for visual pursuit error signaling in the primate retina.J Neurosci. 2015 Apr 1;35(13):5397-408. doi: 10.1523/JNEUROSCI.4369-14.2015. J Neurosci. 2015. PMID: 25834063 Free PMC article.
-
The most numerous ganglion cell type of the mouse retina is a selective feature detector.Proc Natl Acad Sci U S A. 2012 Sep 4;109(36):E2391-8. doi: 10.1073/pnas.1211547109. Epub 2012 Aug 13. Proc Natl Acad Sci U S A. 2012. PMID: 22891316 Free PMC article.
-
Shared and distinct retinal input to the mouse superior colliculus and dorsal lateral geniculate nucleus.J Neurophysiol. 2016 Aug 1;116(2):602-10. doi: 10.1152/jn.00227.2016. Epub 2016 May 11. J Neurophysiol. 2016. PMID: 27169509 Free PMC article.
-
Axonal sodium-channel bands shape the response to electric stimulation in retinal ganglion cells.J Neurophysiol. 2009 Apr;101(4):1972-87. doi: 10.1152/jn.91081.2008. Epub 2009 Feb 4. J Neurophysiol. 2009. PMID: 19193771 Free PMC article.
-
The spatial distribution of glutamatergic inputs to dendrites of retinal ganglion cells.J Comp Neurol. 2008 Sep 10;510(2):221-36. doi: 10.1002/cne.21795. J Comp Neurol. 2008. PMID: 18623177 Free PMC article.
References
-
- Amthor FR, Takahashi ES, Oyster CW. Morphologies of rabbit retinal ganglion cells with complex receptive fields. J Comp Neurol. 1989;280:97–121. - PubMed
-
- Berry MJ. Studying the population code of the retina. In: Stett A, editor. Proceedings MEA meeting 2006. Stuttgart, Germany: BIOPRO Baden-Württemberg GmbH; 2006. pp. 82–85.
-
- Berson DM, Pu M, Famiglietti EV. The zeta cell: a new ganglion cell type in cat retina. J Comp Neurol. 1998;399:269–288. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous