

A diverse and evolutionarily fluid set of microRNAs in Arabidopsis thaliana
- PMID: 17182867
- PMCID: PMC1698448
- DOI: 10.1101/gad.1476406
A diverse and evolutionarily fluid set of microRNAs in Arabidopsis thaliana
Abstract
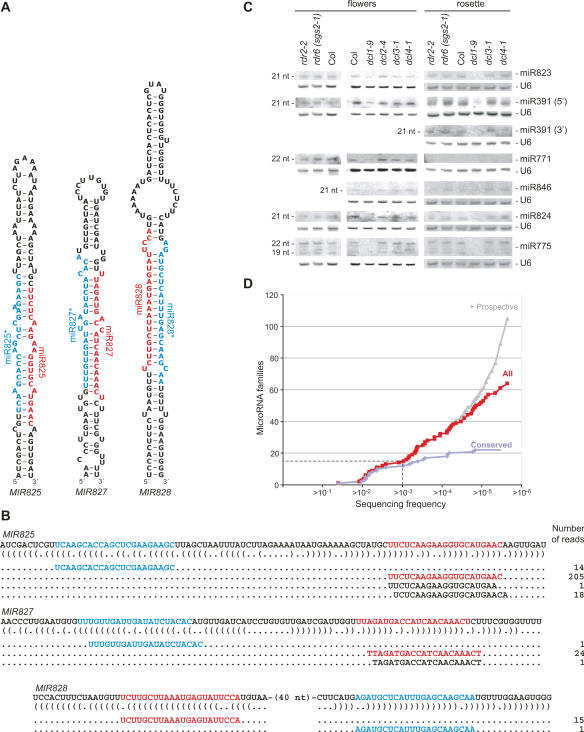
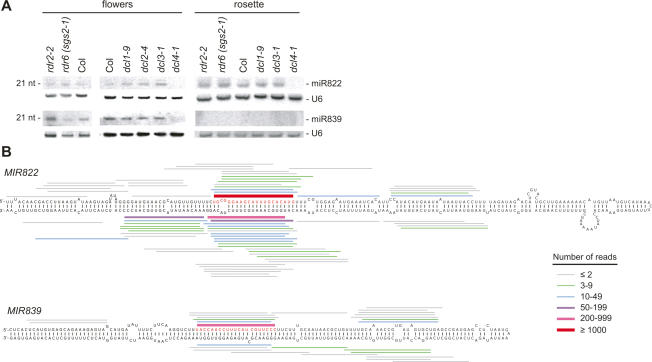
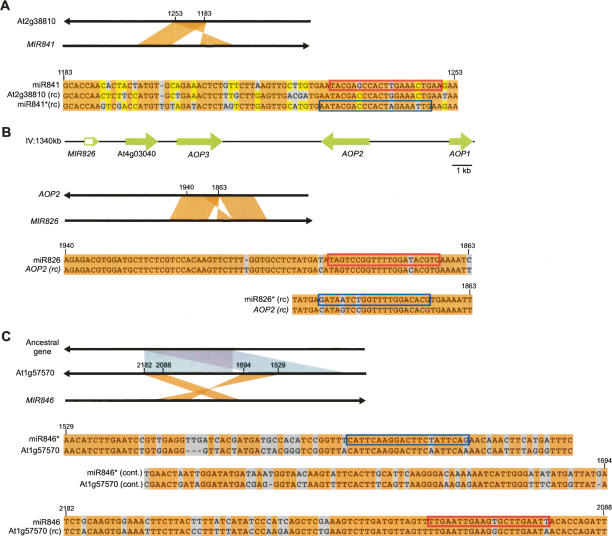
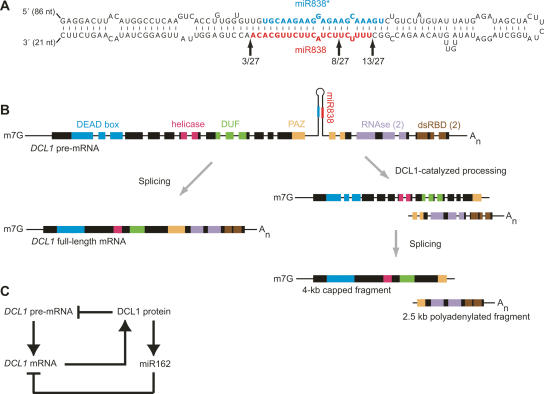
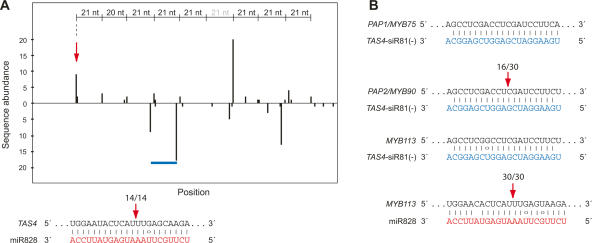
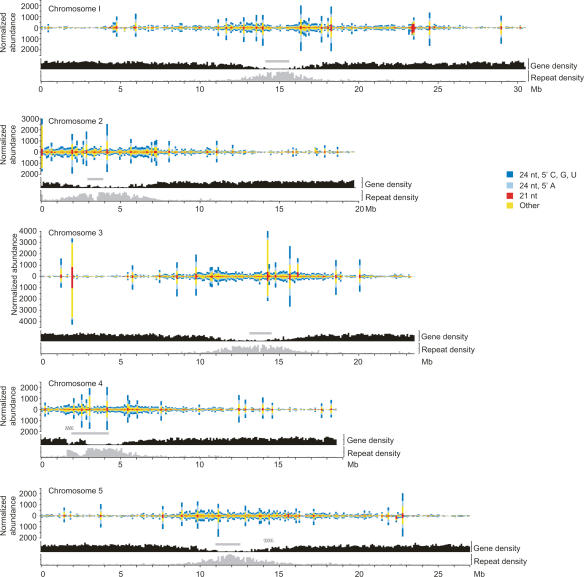
To better understand the diversity of small silencing RNAs expressed in plants, we employed high-throughput pyrosequencing to obtain 887,000 reads corresponding to Arabidopsis thaliana small RNAs. They represented 340,000 unique sequences, a substantially greater diversity than previously obtained in any species. Most of the small RNAs had the properties of heterochromatic small interfering RNAs (siRNAs) associated with DNA silencing in that they were preferentially 24 nucleotides long and mapped to intergenic regions. Their density was greatest in the proximal and distal pericentromeric regions, with only a slightly preferential propensity to match repetitive elements. Also present were 38 newly identified microRNAs (miRNAs) and dozens of other plausible candidates. One miRNA mapped within an intron of DICER-LIKE 1 (DCL1), suggesting a second homeostatic autoregulatory mechanism for DCL1 expression; another defined the phase for siRNAs deriving from a newly identified trans-acting siRNA gene (TAS4); and two depended on DCL4 rather than DCL1 for their accumulation, indicating a second pathway for miRNA biogenesis in plants. More generally, our results revealed the existence of a layer of miRNA-based control beyond that found previously that is evolutionarily much more fluid, employing many newly emergent and diverse miRNAs, each expressed in specialized tissues or at low levels under standard growth conditions.
Figures
References
-
- Adenot, X., Elmayan, T., Lauressergues, D., Boutet, S., Bouche, N., Gasciolli, V., Vaucheret, H. DRB4-dependent TAS3 trans-acting siRNAs control leaf morphology through AGO7. Curr. Biol. 2006;16:927–932. - PubMed
-
- Allen, E., Xie, Z., Gustafson, A.M., Sung, G.H., Spatafora, J.W., Carrington, J.C. Evolution of microRNA genes by inverted duplication of target gene sequences in Arabidopsis thaliana. Nat. Genet. 2004;36:1282–1290. - PubMed
-
- Allen, E., Xie, Z., Gustafson, A.M., Carrington, J.C. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell. 2005;121:207–221. - PubMed
-
- Ambros, V. The functions of animal microRNAs. Nature. 2004;431:350–355. - PubMed
-
- Axtell, M., Jan, C., Rajagopalan, R., Bartel, D.P. A two-hit trigger for siRNA biogenesis in plants. Cell. 2006;127:565–577. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases