

Muscle cachexia is regulated by a p53-PW1/Peg3-dependent pathway
- PMID: 17182869
- PMCID: PMC1698450
- DOI: 10.1101/gad.412606
Muscle cachexia is regulated by a p53-PW1/Peg3-dependent pathway
Abstract
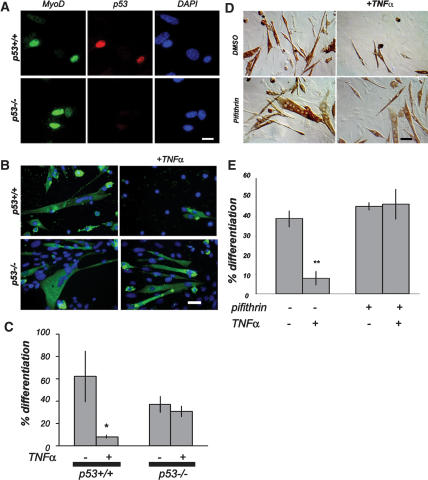
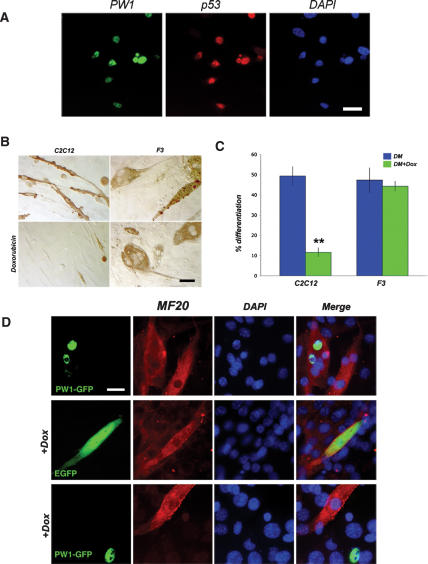
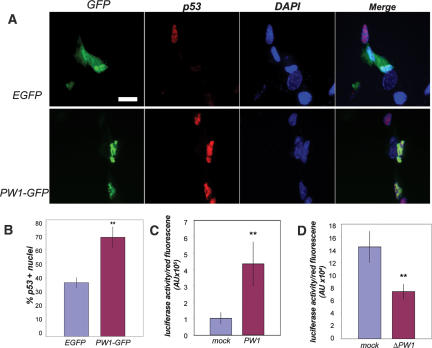
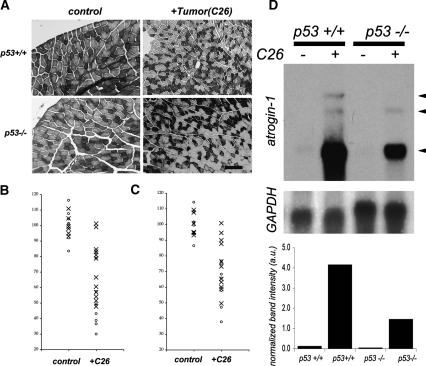
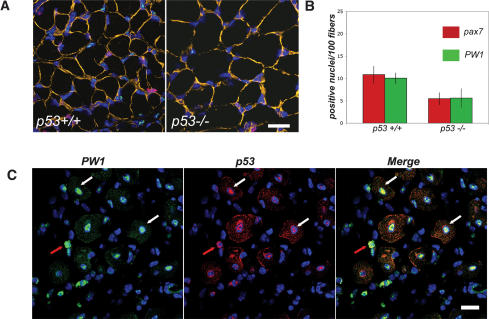
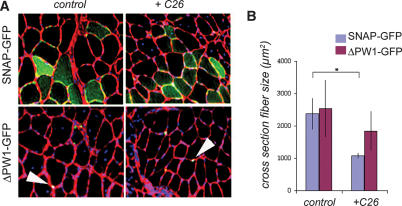
Muscle wasting (cachexia) is an incurable complication associated with chronic infection and cancers that leads to an overall poor prognosis for recovery. Tumor necrosis factor-alpha (TNFalpha) is a key inflammatory cytokine associated with cachexia. TNFalpha inhibits myogenic differentiation and skeletal muscle regeneration through downstream effectors of the p53 cell death pathway including PW1/Peg3, bax, and caspases. We report that p53 is required for the TNFalpha-mediated inhibition of myogenesis in vitro and contributes to muscle wasting in response to tumor load in vivo. We further demonstrate that PW1 and p53 participate in a positive feedback regulatory loop in vitro. Consistent with this observation, we find that the number of PW1-expressing stem cells in skeletal muscle declines significantly in p53 nullizygous mice. Furthermore, gene transfer of a dominant-negative form of PW1 into muscle tissue in vivo blocks myofiber atrophy in response to tumor load. Taken together, these results show a novel role for p53 in mediating muscle stem cell behavior and muscle atrophy, and point to new targets for the therapeutic treatment of muscle wasting.
Figures
References
-
- Acharyya, S., Butchbach, M.E., Sahenk, Z., Wang, H., Saji, M., Carathers, M., Ringel, M.D., Skipworth, R.J., Fearon, K.C., Hollingsworth, M.A., et al. Dystrophin glycoprotein complex dysfunction: A regulatory link between muscular dystrophy and cancer cachexia. Cancer Cell. 2005;8:421–432. - PubMed
-
- Bodine, S.C., Stitt, T.N., Gonzalez, M., Kline, W.O., Stover, G.L., Bauerlein, R., Zlotchenko, E., Scrimgeour, A., Lawrence, J.C., Glass, D.J., et al. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat. Cell Biol. 2001;3:1014–1019. - PubMed
-
- Cai, D., Frantz, J.D., Tawa, N.E., Jr., Melendez, P.A., Oh, B.C., Lidov, H.G., Hasselgren, P.O., Frontera, W.R., Lee, J., Glass, D.J., et al. IKKβ/NF-κB activation causes severe muscle wasting in mice. Cell. 2004;119:285–298. - PubMed
-
- Charge, S.B., Rudnicki, M.A. Cellular and molecular regulation of muscle regeneration. Physiol. Rev. 2004;84:209–238. - PubMed
-
- Chin, L., Artandi, S.E., Shen, Q., Tam, A., Lee, S.L., Gottlieb, G.J., Greider, C.W., DePinho, R.A. p53 deficiency rescues the adverse effects of telomere loss and cooperates with telomere dysfunction to accelerate carcinogenesis. Cell. 1999;97:527–538. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous