

Chemokine receptor expression by neural progenitor cells in neurogenic regions of mouse brain
- PMID: 17183554
- PMCID: PMC2758702
- DOI: 10.1002/cne.21229
Chemokine receptor expression by neural progenitor cells in neurogenic regions of mouse brain
Abstract
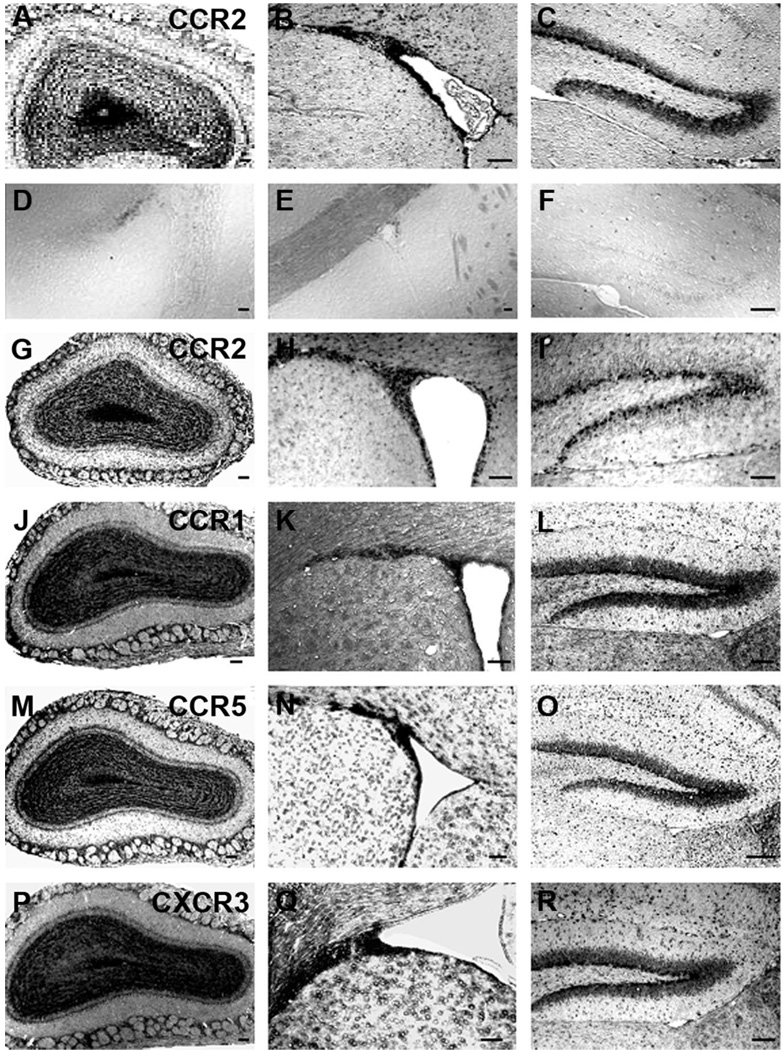
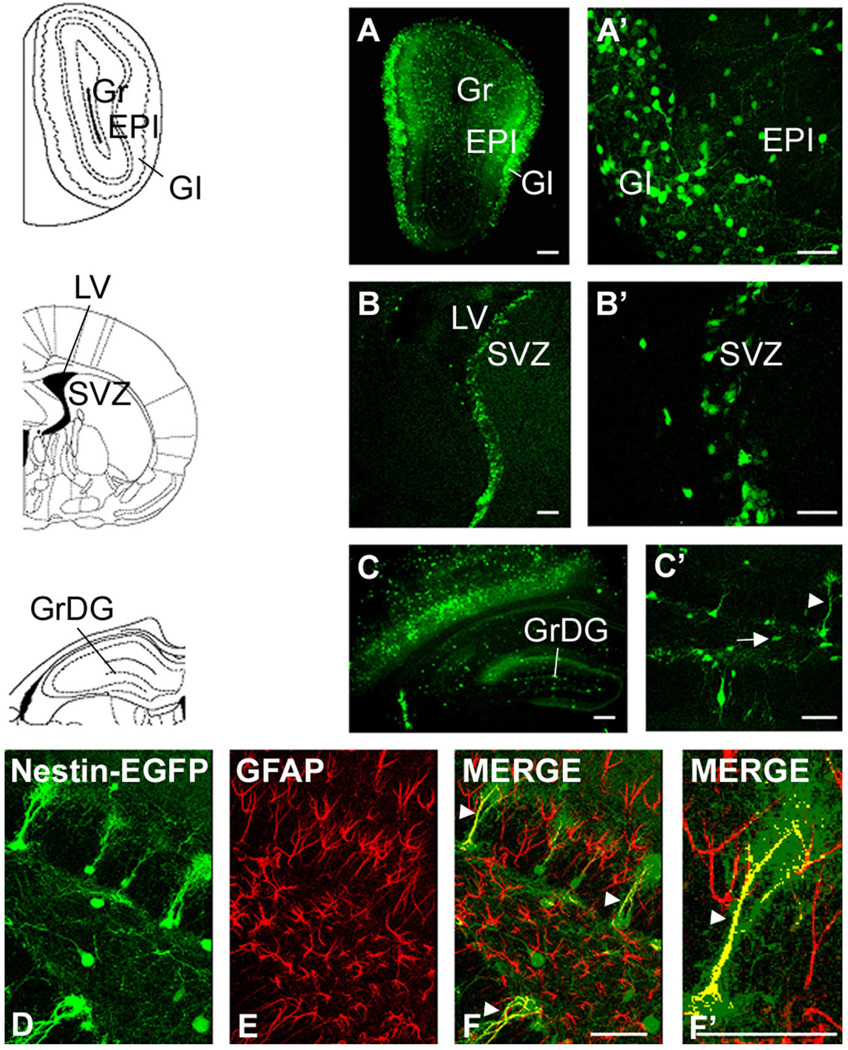
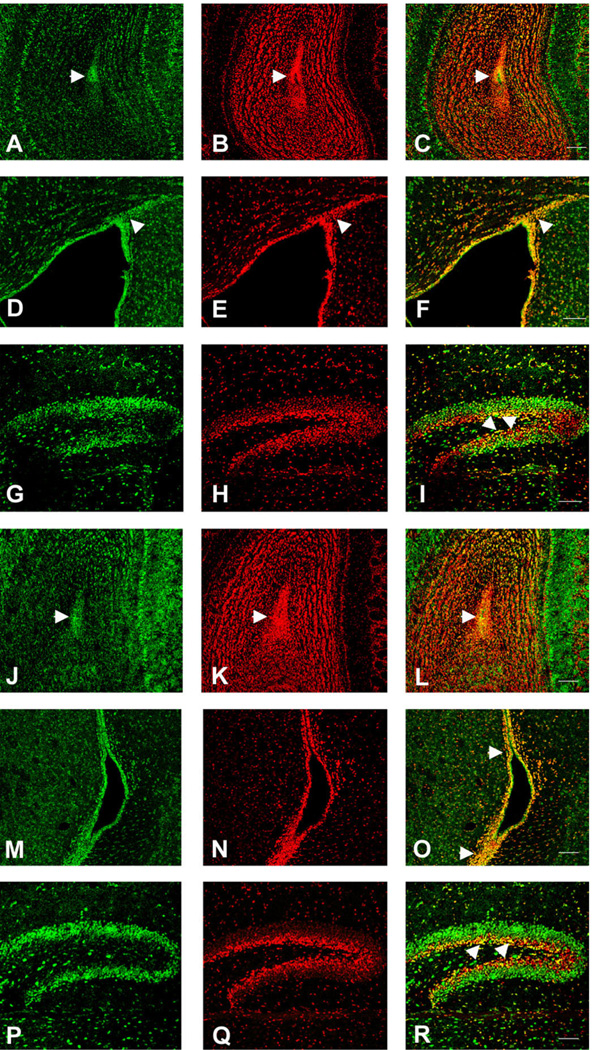
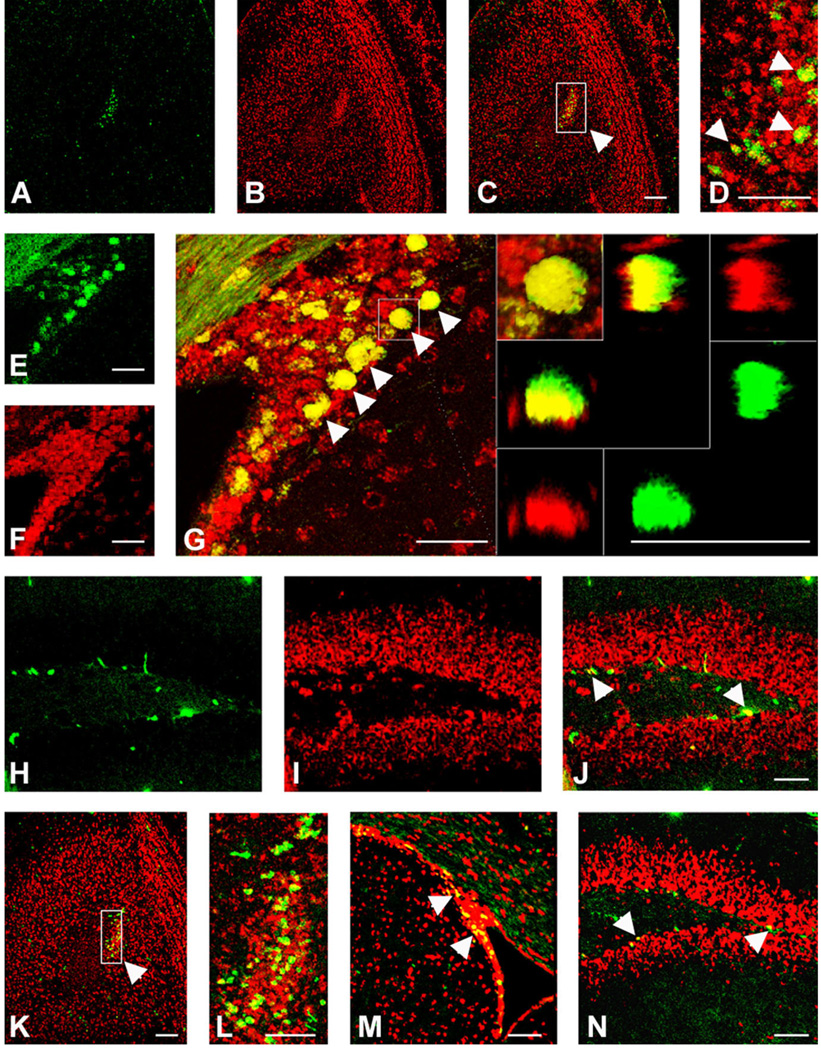
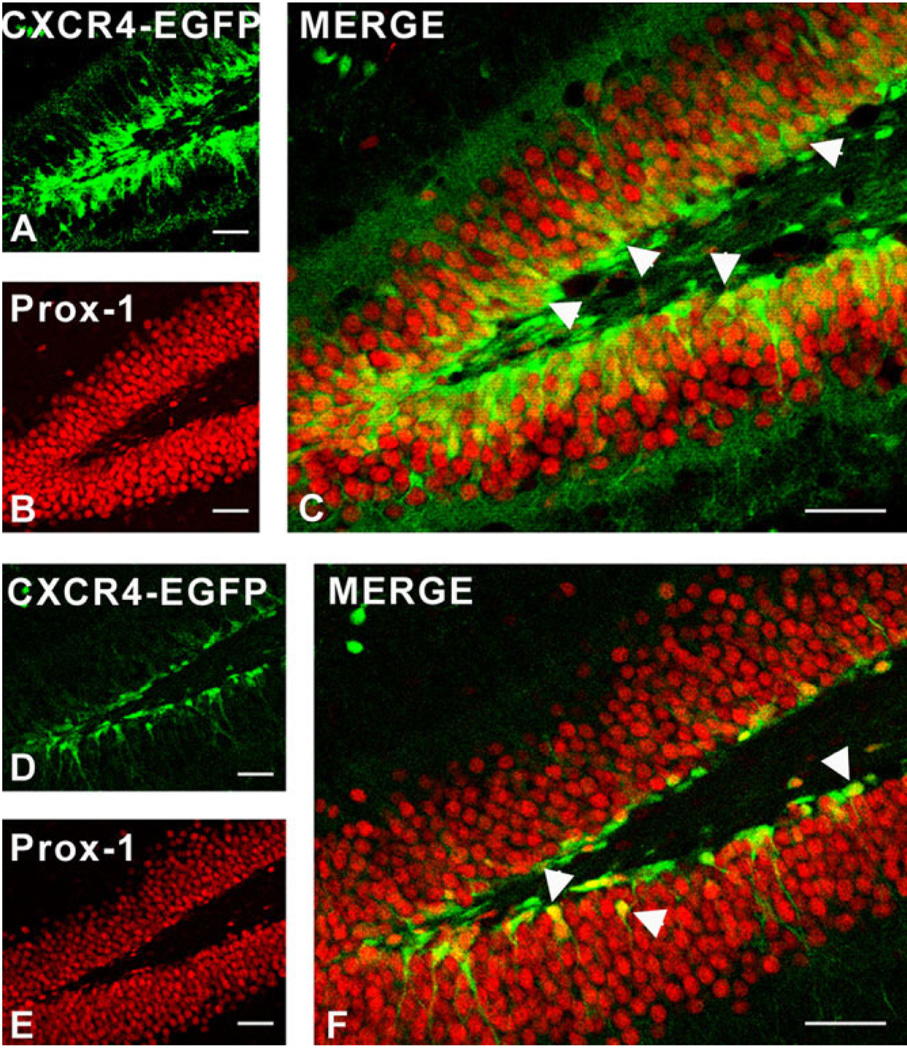
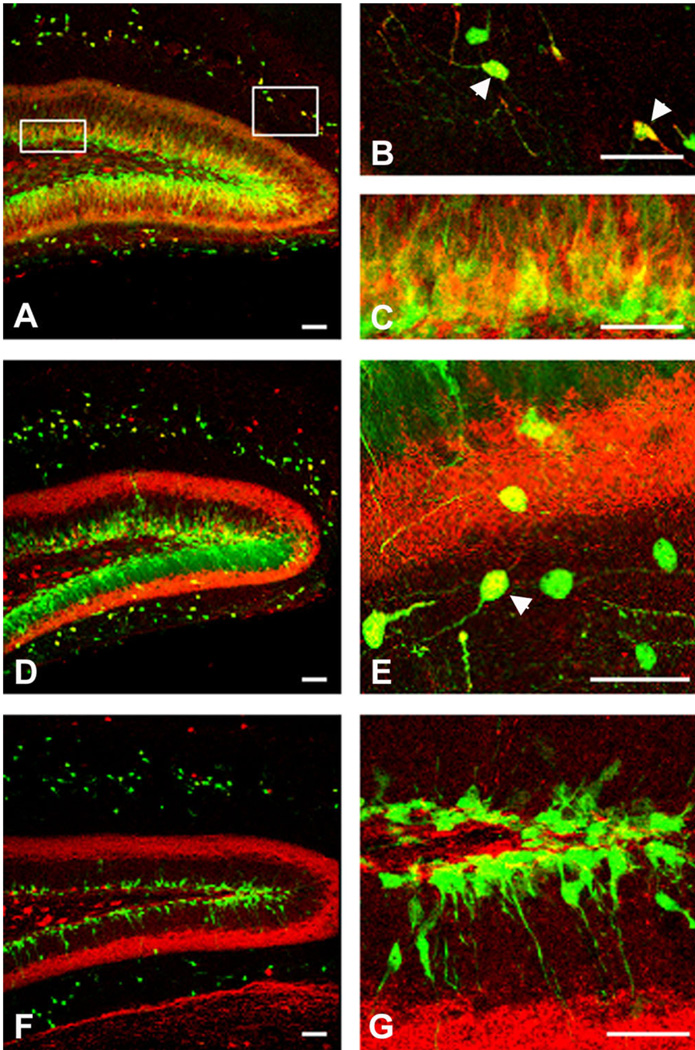
We previously demonstrated that chemokine receptors are expressed by neural progenitors grown as cultured neurospheres. To examine the significance of these findings for neural progenitor function in vivo, we investigated whether chemokine receptors were expressed by cells having the characteristics of neural progenitors in neurogenic regions of the postnatal brain. Using in situ hybridization we demonstrated the expression of CCR1, CCR2, CCR5, CXCR3, and CXCR4 chemokine receptors by cells in the dentate gyrus (DG), subventricular zone of the lateral ventricle, and olfactory bulb. The pattern of expression for all of these receptors was similar, including regions where neural progenitors normally reside. In addition, we attempted to colocalize chemokine receptors with markers for neural progenitors. In order to do this we used nestin-EGFP and TLX-LacZ transgenic mice, as well as labeling for Ki67, a marker for dividing cells. In all three areas of the brain we demonstrated colocalization of chemokine receptors with these three markers in populations of cells. Expression of chemokine receptors by neural progenitors was further confirmed using CXCR4-EGFP BAC transgenic mice. Expression of CXCR4 in the DG included cells that expressed nestin and GFAP as well as cells that appeared to be immature granule neurons expressing PSA-NCAM, calretinin, and Prox-1. CXCR4-expressing cells in the DG were found in close proximity to immature granule neurons that expressed the chemokine SDF-1/CXCL12. Cells expressing CXCR4 frequently coexpressed CCR2 receptors. These data support the hypothesis that chemokine receptors are important in regulating the migration of progenitor cells in postnatal brain.
Figures














References
-
- Aboody KS, Brown A, Rainov NG, Bower KA, Liu S, Yang W, Small JE, Herrlinger U, Ourednik V, Black PM, Breakefield XO, Snyder EY. Neural stem cells display extensive tropism for pathology in adult brain: evidence from intracranial gliomas. Proc Natl Acad Sci U S A. 2000;97:12846–12851. - PMC - PubMed
-
- Abrous DN, Koehl M, Le Moal M. Adult neurogenesis: from precursors to network and physiology. Physiol Rev. 2005;85:523–569. - PubMed
-
- Altman J, Das GD. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J Comp Neurol. 1965;124:319–336. - PubMed
-
- Alvarez-Buylla A, Lim DA. For the long run: maintaining germinal niches in the adult brain. Neuron. 2004;41:683–686. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
