

Acidic residues of yeast frataxin have an essential role in Fe-S cluster assembly
- PMID: 17186026
- PMCID: PMC1796776
- DOI: 10.1038/sj.embor.7400881
Acidic residues of yeast frataxin have an essential role in Fe-S cluster assembly
Abstract
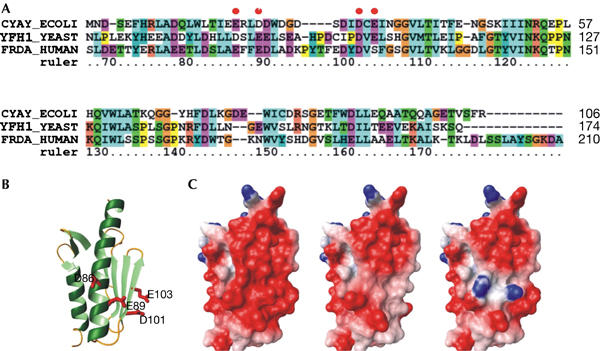
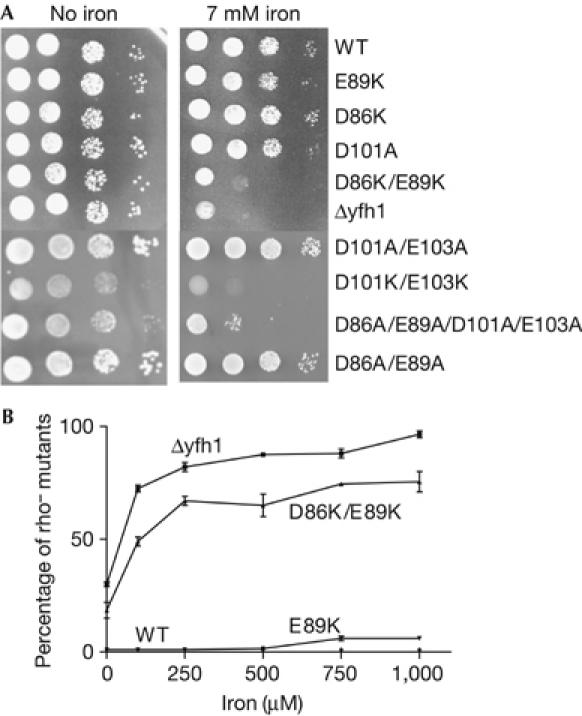
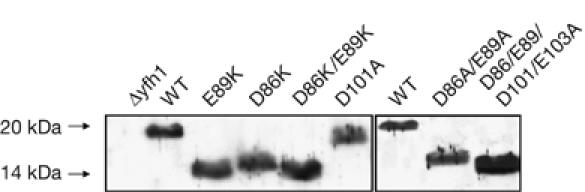
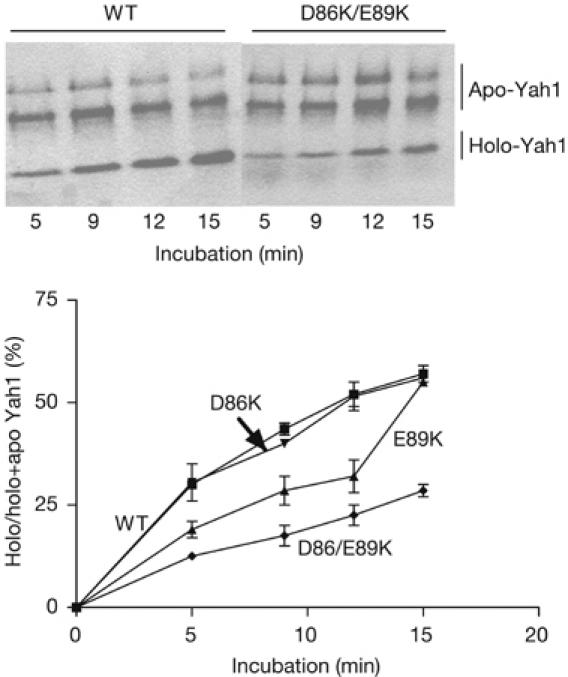
Friedreich ataxia is caused by decreased levels of frataxin, a mitochondrial acidic protein that is assumed to act as chaperone in the assembly of Fe-S clusters on the scaffold Isu protein. Frataxin has the in vitro capacity to form iron-loaded multimers, which also suggests an iron storage function. It has been reported that alanine substitution of residues in an acidic ridge of yeast frataxin (Yfh1) elicits loss of iron binding in vitro but has no effect on Fe-S cluster synthesis in vivo. Here, we show that a marked change in the electrostatic properties of a specific region of Yfh1 surface - by substituting two or four acidic residues by lysine or alanine, respectively - impairs Fe-S cluster assembly, weakens the interaction between Yfh1 and Isu1, and increases oxidative damage. Therefore, the acidic ridge is essential for the Yfh1 function and is likely to be involved in iron-mediated protein-protein interactions.
Figures
References
-
- Adinolfi S, Trifuoggi M, Politou AS, Martin S, Pastore A (2002) A structural approach to understanding the iron-binding properties of phylogenetically different frataxins. Hum Mol Genet 11: 1865–1877 - PubMed
-
- Babcock M, de Silva D, Oaks R, Davis-Kaplan S, Jiralerspong S, Montermini L, Pandolfo M, Kaplan J (1997) Regulation of mitochondrial iron accumulation by Yfh1p, a putative homolog of frataxin. Science 276: 1709–1712 - PubMed
-
- Branda SS, Cavadini P, Adamec J, Kalousek F, Taroni F, Isaya G (1999) Yeast and human frataxin are processed to mature form in two sequential steps by the mitochondrial processing peptidase. J Biol Chem 274: 22763–22769 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
