

The role of cholesterol and sphingolipids in chemokine receptor function and HIV-1 envelope glycoprotein-mediated fusion
- PMID: 17187670
- PMCID: PMC1769366
- DOI: 10.1186/1743-422X-3-104
The role of cholesterol and sphingolipids in chemokine receptor function and HIV-1 envelope glycoprotein-mediated fusion
Abstract
Background: HIV-1 entry into cells is a multifaceted process involving target cell CD4 and the chemokine receptors, CXCR4 or CCR5. The lipid composition of the host cell plays a significant role in the HIV fusion process as it orchestrates the appropriate disposition of CD4 and co-receptors required for HIV-1 envelope glycoprotein (Env)-mediated fusion. The cell membrane is primarily composed of sphingolipids and cholesterol. The effects of lipid modulation on CD4 disposition in the membrane and their role in HIV-1 entry have extensively been studied. To focus on the role of lipid composition on chemokine receptor function, we have by-passed the CD4 requirement for HIV-1 Env-mediated fusion by using a CD4-independent strain of HIV-1 Env.
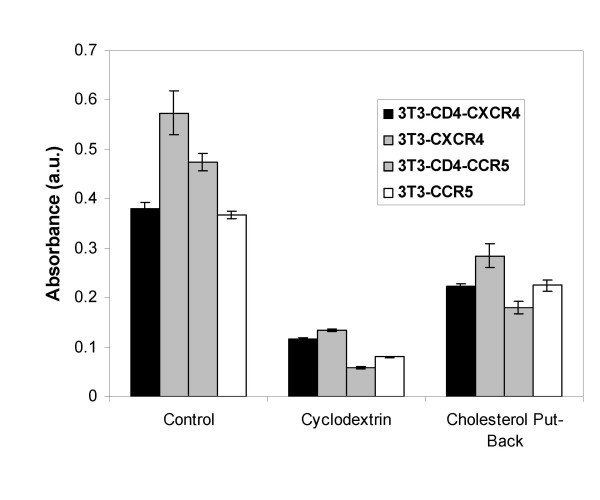
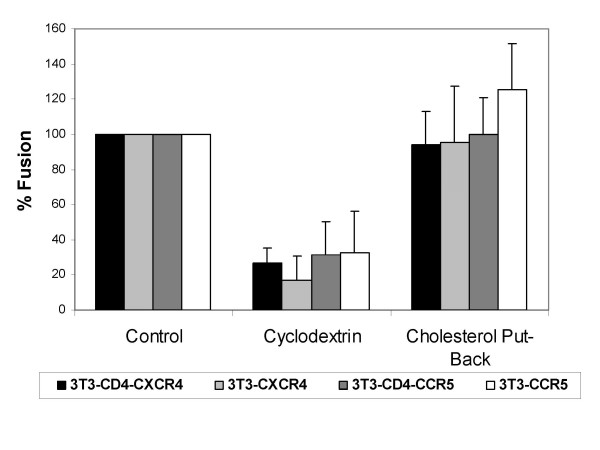
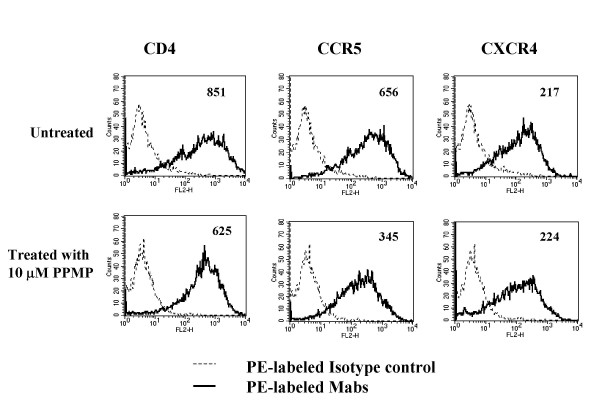
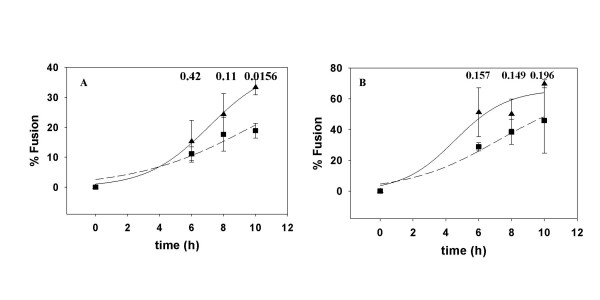
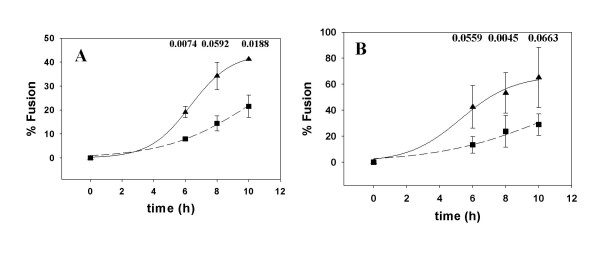
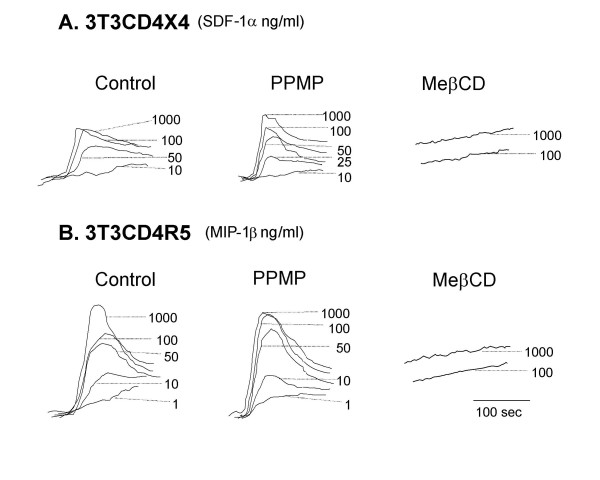
Results: Cell fusion mediated by a CD4-independent strain of HIV-1 Env was monitored by observing dye transfer between Env-expressing cells and NIH3T3 cells bearing CXCR4 or CCR5 in the presence or absence of CD4. Chemokine receptor signaling was assessed by monitoring changes in intracellular [Ca2+] mobilization induced by CCR5 or CXCR4 ligand. To modulate target membrane cholesterol or sphingolipids we used Methyl-beta-cyclodextrin (MbetaCD) or 1-phenyl-2-hexadecanoylamino-3-morpholino-1-propanol (PPMP), respectively. Treatment of the target cells with these agents did not change the levels of CD4 or CXCR4, but reduced levels of CCR5 on the cell surface. Chemokine receptor signalling was inhibited by cholesterol removal but not by treatment with PPMP. HIV-1 Env mediated fusion was inhibited by >50% by cholesterol removal. Overall, PPMP treatment appeared to slow down the rates of CD4-independent HIV-1 Env-mediated Fusion. However, in the case of CXCR4-dependent fusion, the differences between untreated and PPMP-treated cells did not appear to be significant.
Conclusion: Although modulation of cholesterol and sphingolipids has similar effects on CD4-dependent HIV-1 Env-mediated fusion, sphingolipid modulation had little effect on CD4-independent HIV-1 Env-mediated fusion. Chemokine receptor function remained intact following treatment of cells with PPMP. Therefore such treatment may be considered a more suitable agent to inhibit CD4 dependent HIV-1 infection.
Figures
References
-
- Moulard M, Decroly E. Maturation of HIV envelope glycoprotein precursors by cellular endoproteases. Biochim Biophys Acta. 2000;1469:121–132. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
