

Collagen reorganization at the tumor-stromal interface facilitates local invasion
- PMID: 17190588
- PMCID: PMC1781458
- DOI: 10.1186/1741-7015-4-38
Collagen reorganization at the tumor-stromal interface facilitates local invasion
Abstract
Background: Stromal-epithelial interactions are of particular significance in breast tissue as misregulation of these interactions can promote tumorigenesis and invasion. Moreover, collagen-dense breast tissue increases the risk of breast carcinoma, although the relationship between collagen density and tumorigenesis is not well understood. As little is known about epithelial-stromal interactions in vivo, it is necessary to visualize the stroma surrounding normal epithelium and mammary tumors in intact tissues to better understand how matrix organization, density, and composition affect tumor formation and progression.
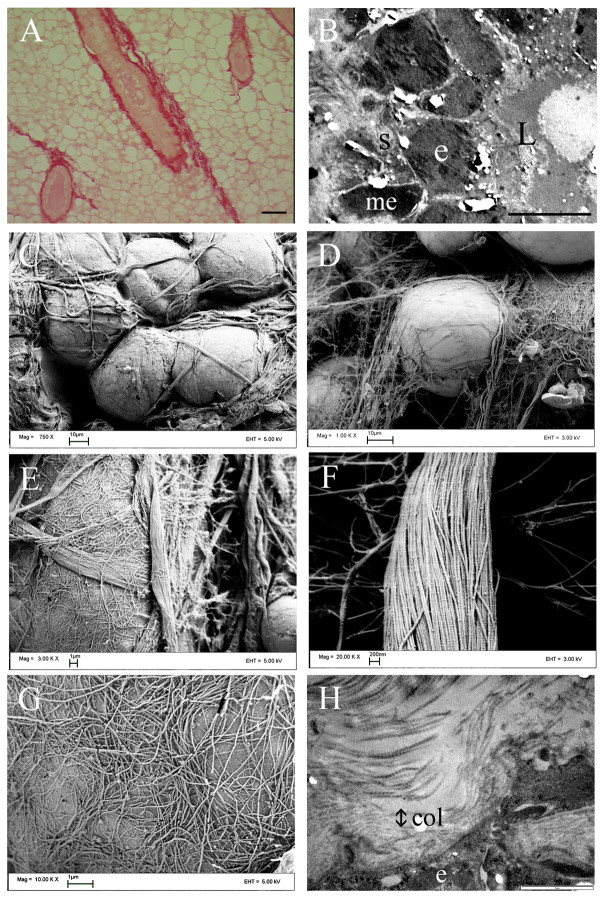
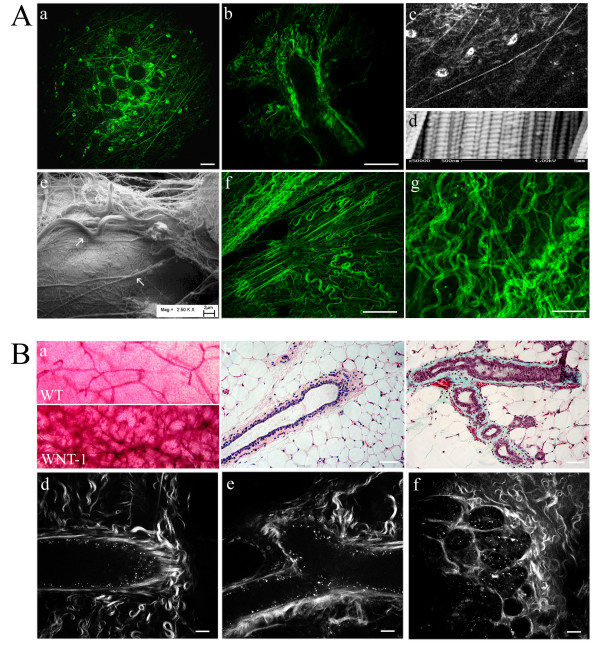
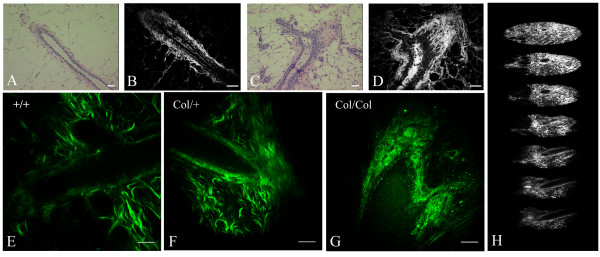
Methods: Epithelial-stromal interactions in normal mammary glands, mammary tumors, and tumor explants in three-dimensional culture were studied with histology, electron microscopy, and nonlinear optical imaging methodologies. Imaging of the tumor-stromal interface in live tumor tissue ex vivo was performed with multiphoton laser-scanning microscopy (MPLSM) to generate multiphoton excitation (MPE) of endogenous fluorophores and second harmonic generation (SHG) to image stromal collagen.
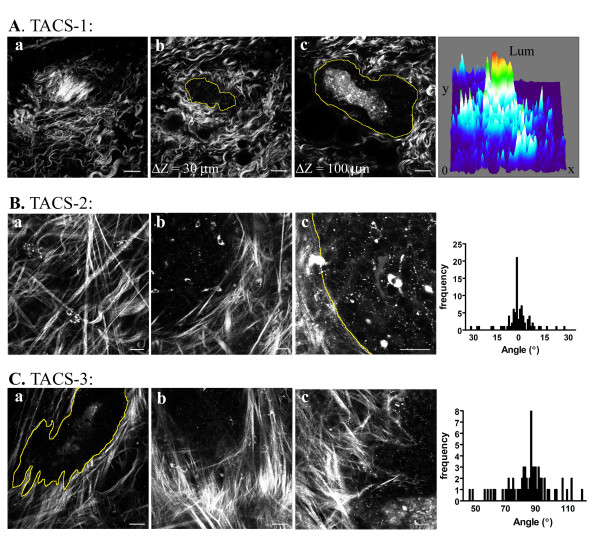
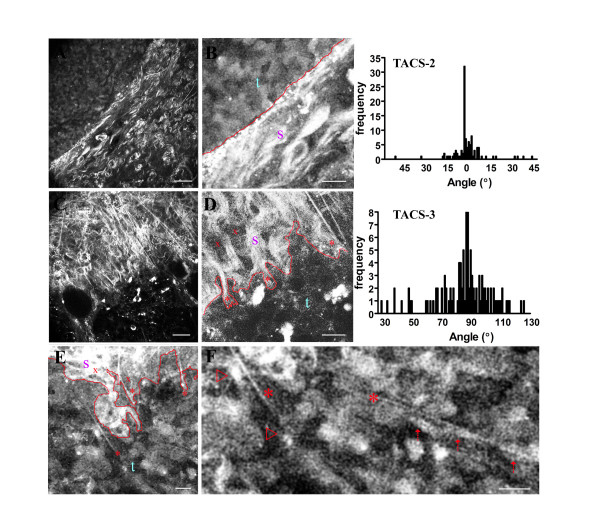
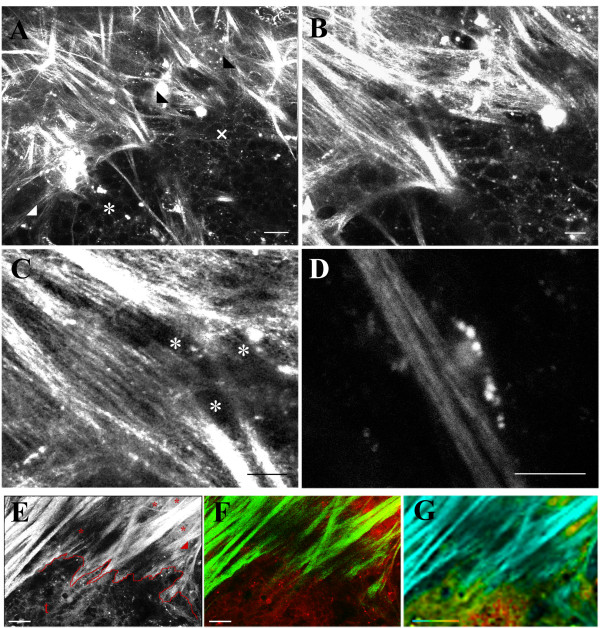
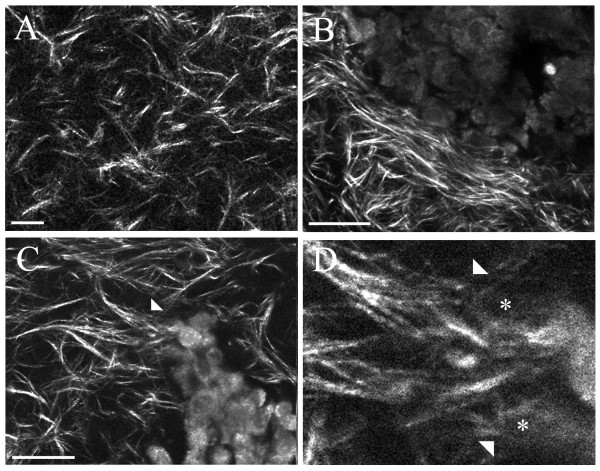
Results: We used both laser-scanning multiphoton and second harmonic generation microscopy to determine the organization of specific collagen structures around ducts and tumors in intact, unfixed and unsectioned mammary glands. Local alterations in collagen density were clearly seen, allowing us to obtain three-dimensional information regarding the organization of the mammary stroma, such as radiating collagen fibers that could not have been obtained using classical histological techniques. Moreover, we observed and defined three tumor-associated collagen signatures (TACS) that provide novel markers to locate and characterize tumors. In particular, local cell invasion was found predominantly to be oriented along certain aligned collagen fibers, suggesting that radial alignment of collagen fibers relative to tumors facilitates invasion. Consistent with this observation, primary tumor explants cultured in a randomly organized collagen matrix realigned the collagen fibers, allowing individual tumor cells to migrate out along radially aligned fibers.
Conclusion: The presentation of these tumor-associated collagen signatures allowed us to identify pre-palpable tumors and see cells at the tumor-stromal boundary invading into the stroma along radially aligned collagen fibers. As such, TACS should provide indications that a tumor is, or could become, invasive, and may serve as part of a strategy to help identify and characterize breast tumors in animal and human tissues.
Figures
References
-
- Keely P, Fong A, Zutter M, Santoro S. Alteration of collagen-dependent adhesion, motility, and morphogenesis by the expression of antisense α2 integrin mRNA in mammary cells. J Cell Science. 1995;108:595–607. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
