

Activity-dependent neurotransmitter-receptor matching at the neuromuscular junction
- PMID: 17190810
- PMCID: PMC1749326
- DOI: 10.1073/pnas.0607450104
Activity-dependent neurotransmitter-receptor matching at the neuromuscular junction
Abstract
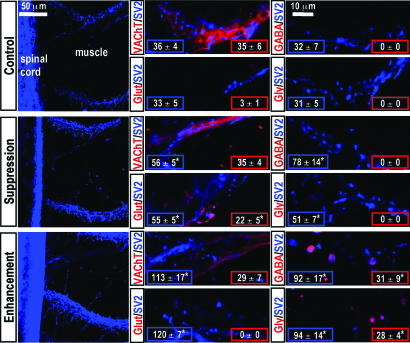
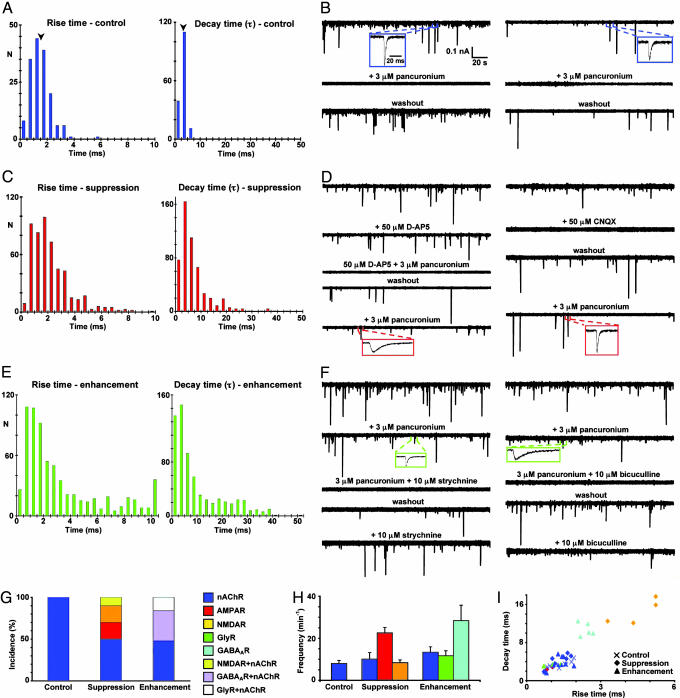
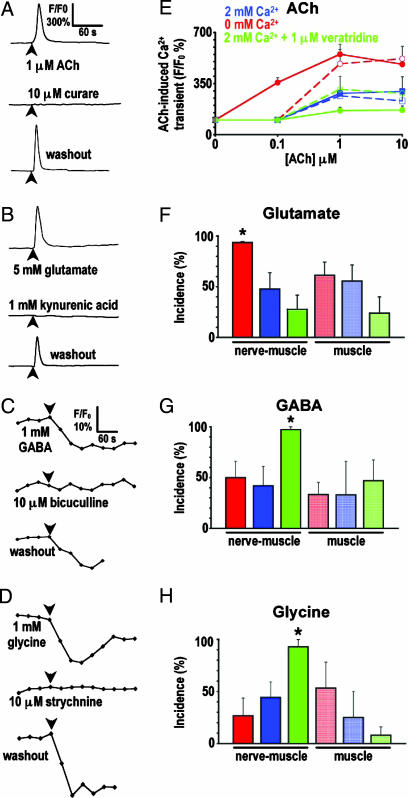
Signaling in the nervous system requires matching of neurotransmitter receptors with cognate neurotransmitters at synapses. The vertebrate neuromuscular junction is the best studied cholinergic synapse, but the mechanisms by which acetylcholine is matched with acetylcholine receptors are not fully understood. Because alterations in neuronal calcium spike activity alter transmitter specification in embryonic spinal neurons, we hypothesized that receptor expression in postsynaptic cells follows changes in transmitter expression to achieve this specific match. We find that embryonic vertebrate striated muscle cells normally express receptors for glutamate, GABA, and glycine as well as for acetylcholine. As maturation progresses, acetylcholine receptor expression prevails. Receptor selection is altered when early neuronal calcium-dependent activity is perturbed, and remaining receptor populations parallel changes in transmitter phenotype. In these cases, glutamatergic, GABAergic, and glycinergic synaptic currents are recorded from muscle cells, demonstrating that activity regulates matching of transmitters and their receptors in the assembly of functional synapses.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
