

Phytotoxicity and innate immune responses induced by Nep1-like proteins
- PMID: 17194768
- PMCID: PMC1785393
- DOI: 10.1105/tpc.106.044180
Phytotoxicity and innate immune responses induced by Nep1-like proteins
Abstract
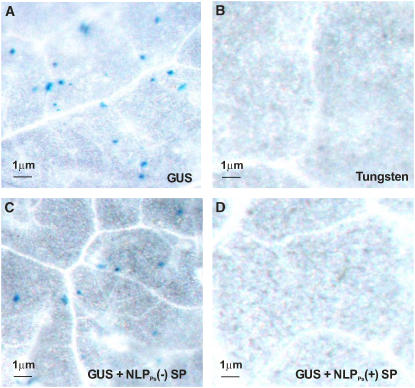
We show that oomycete-derived Nep1 (for necrosis and ethylene-inducing peptide1)-like proteins (NLPs) trigger a comprehensive immune response in Arabidopsis thaliana, comprising posttranslational activation of mitogen-activated protein kinase activity, deposition of callose, production of nitric oxide, reactive oxygen intermediates, ethylene, and the phytoalexin camalexin, as well as cell death. Transcript profiling experiments revealed that NLPs trigger extensive reprogramming of the Arabidopsis transcriptome closely resembling that evoked by bacteria-derived flagellin. NLP-induced cell death is an active, light-dependent process requiring HSP90 but not caspase activity, salicylic acid, jasmonic acid, ethylene, or functional SGT1a/SGT1b. Studies on animal, yeast, moss, and plant cells revealed that sensitivity to NLPs is not a general characteristic of phospholipid bilayer systems but appears to be restricted to dicot plants. NLP-induced cell death does not require an intact plant cell wall, and ectopic expression of NLP in dicot plants resulted in cell death only when the protein was delivered to the apoplast. Our findings strongly suggest that NLP-induced necrosis requires interaction with a target site that is unique to the extracytoplasmic side of dicot plant plasma membranes. We propose that NLPs play dual roles in plant pathogen interactions as toxin-like virulence factors and as triggers of plant innate immune responses.
Figures








Similar articles
-
The Nicotiana benthamiana mitogen-activated protein kinase cascade and WRKY transcription factor participate in Nep1(Mo)-triggered plant responses.Mol Plant Microbe Interact. 2012 Dec;25(12):1639-53. doi: 10.1094/MPMI-11-11-0293. Mol Plant Microbe Interact. 2012. PMID: 22835275
-
Nep1-like proteins from three kingdoms of life act as a microbe-associated molecular pattern in Arabidopsis.Proc Natl Acad Sci U S A. 2014 Nov 25;111(47):16955-60. doi: 10.1073/pnas.1410031111. Epub 2014 Nov 3. Proc Natl Acad Sci U S A. 2014. PMID: 25368167 Free PMC article.
-
NPP1, a Phytophthora-associated trigger of plant defense in parsley and Arabidopsis.Plant J. 2002 Nov;32(3):375-90. doi: 10.1046/j.1365-313x.2002.01454.x. Plant J. 2002. PMID: 12410815
-
Activity and Phylogenetics of the Broadly Occurring Family of Microbial Nep1-Like Proteins.Annu Rev Phytopathol. 2019 Aug 25;57:367-386. doi: 10.1146/annurev-phyto-082718-100054. Epub 2019 Jul 5. Annu Rev Phytopathol. 2019. PMID: 31283435 Review.
-
Disruption of plant plasma membrane by Nep1-like proteins in pathogen-plant interactions.New Phytol. 2023 Feb;237(3):746-750. doi: 10.1111/nph.18524. Epub 2022 Nov 29. New Phytol. 2023. PMID: 36210522 Free PMC article. Review.
Cited by
-
The role of vacuolar processing enzyme (VPE) from Nicotiana benthamiana in the elicitor-triggered hypersensitive response and stomatal closure.J Exp Bot. 2010 Aug;61(13):3799-812. doi: 10.1093/jxb/erq189. Epub 2010 Jul 5. J Exp Bot. 2010. PMID: 20603283 Free PMC article.
-
Functional Identification of Novel Cell Death-inducing Effector Proteins from Magnaporthe oryzae.Rice (N Y). 2019 Aug 6;12(1):59. doi: 10.1186/s12284-019-0312-z. Rice (N Y). 2019. PMID: 31388773 Free PMC article.
-
An RXLR effector secreted by Phytophthora parasitica is a virulence factor and triggers cell death in various plants.Mol Plant Pathol. 2019 Mar;20(3):356-371. doi: 10.1111/mpp.12760. Epub 2018 Nov 22. Mol Plant Pathol. 2019. PMID: 30320960 Free PMC article.
-
The Secreted Ribonuclease SRE1 Contributes to Setosphaeria turcica Virulence and Activates Plant Immunity.Front Microbiol. 2022 Jul 8;13:941991. doi: 10.3389/fmicb.2022.941991. eCollection 2022. Front Microbiol. 2022. PMID: 35875548 Free PMC article.
-
Global gene expression of Poncirus trifoliata, Citrus sunki and their hybrids under infection of Phytophthora parasitica.BMC Genomics. 2011 Jan 17;12:39. doi: 10.1186/1471-2164-12-39. BMC Genomics. 2011. PMID: 21241495 Free PMC article.
References
-
- Akira, S., Uematsu, S., and Takeuchi, O. (2006). Pathogen recognition and innate immunity. Cell 124 783–801. - PubMed
-
- Alfano, J.R., and Collmer, A. (2004). Type III secretion system effector proteins: Double agents in bacterial disease and plant defense. Annu. Rev. Phytopathol. 42 385–414. - PubMed
-
- Amsellem, Z., Cohen, B.A., and Gressel, J. (2002). Engineering hypervirulence in a mycoherbicidal fungus for efficient weed control. Nat. Biotechnol. 20 1035–1039. - PubMed
-
- Asai, T., Tena, G., Plotnikova, J., Willmann, M.R., Chiu, W.L., Gomez-Gomez, L., Boller, T., Ausubel, F.M., and Sheen, J. (2002). MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 415 977–983. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
