

Cyclin and cyclin-dependent kinase substrate requirements for preventing rereplication reveal the need for concomitant activation and inhibition
- PMID: 17194775
- PMCID: PMC1840059
- DOI: 10.1534/genetics.106.068213
Cyclin and cyclin-dependent kinase substrate requirements for preventing rereplication reveal the need for concomitant activation and inhibition
Abstract
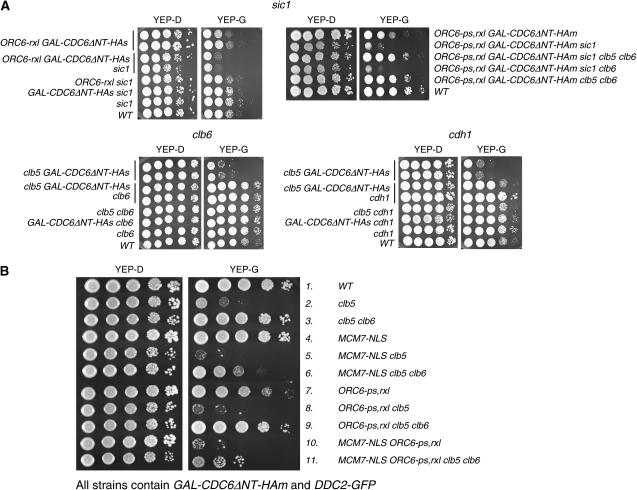
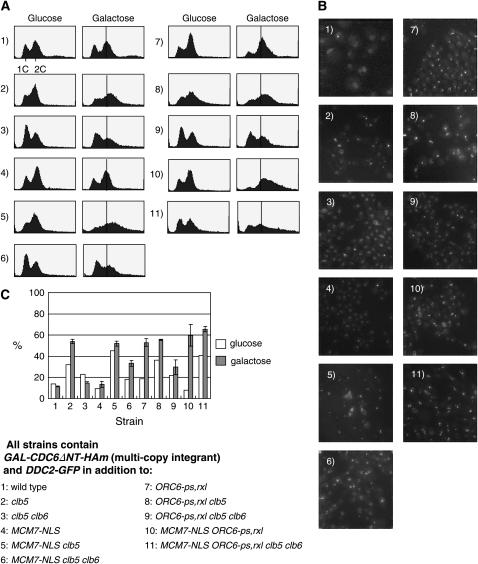
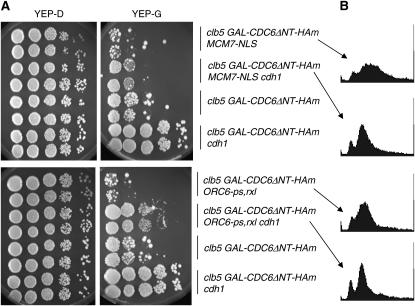
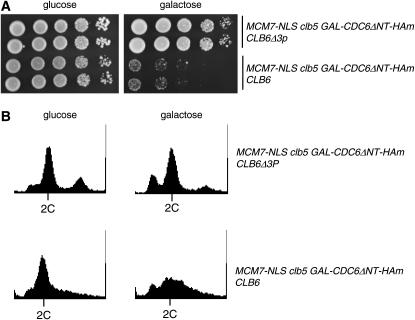
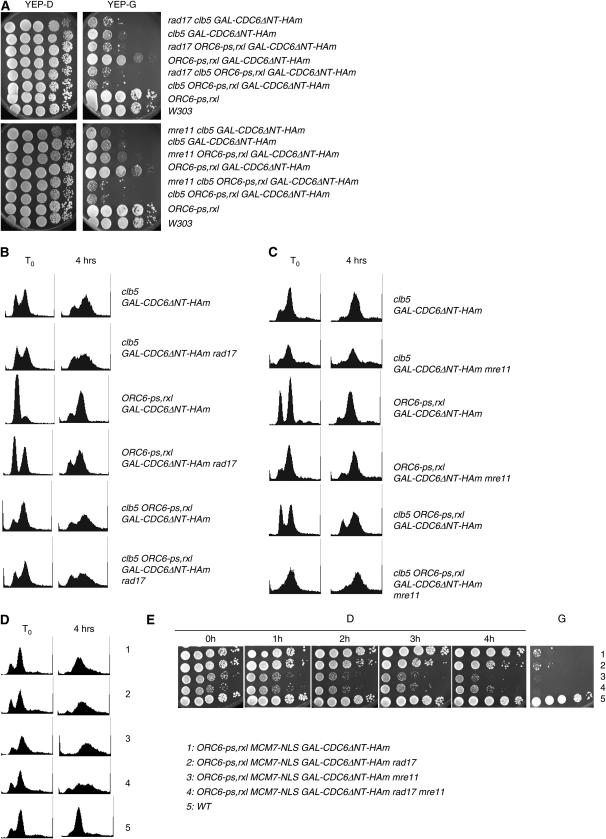
DNA replication initiation in S. cerevisiae is promoted by B-type cyclin-dependent kinase (Cdk) activity. In addition, once-per-cell-cycle replication is enforced by cyclin-Cdk-dependent phosphorylation of the prereplicative complex (pre-RC) components Mcm2-7, Cdc6, and Orc1-6. Several of these controls must be simultaneously blocked by mutation to obtain rereplication. We looked for but did not obtain strong evidence for cyclin specificity in the use of different mechanisms to control rereplication: both the S-phase cyclin Clb5 and the mitotic cyclins Clb1-4 were inferred to be capable of imposing ORC-based and MCM-based controls. We found evidence that the S-phase cyclin Clb6 could promote initiation of replication without blocking reinitiation, and this activity was highly toxic when the ability of other cyclins to block reinitiation was prevented by mutation. The failure of Clb6 to regulate reinitiation was due to rapid Clb6 proteolysis, since this toxic activity of Clb6 was lost when Clb6 was stabilized by mutation. Clb6-dependent toxicity is also relieved when early accumulation of mitotic cyclins is allowed to impose rereplication controls. Cell-cycle timing of rereplication control is crucial: sufficient rereplication block activity must be available as soon as firing begins. DNA rereplication induces DNA damage, and when rereplication controls are compromised, the DNA damage checkpoint factors Mre11 and Rad17 provide additional mechanisms that maintain viability and also prevent further rereplication, and this probably contributes to genome stability.
Figures
Similar articles
-
S-phase-promoting cyclin-dependent kinases prevent re-replication by inhibiting the transition of replication origins to a pre-replicative state.Curr Biol. 1995 Nov 1;5(11):1257-69. doi: 10.1016/s0960-9822(95)00252-1. Curr Biol. 1995. PMID: 8574583
-
CLB5 and CLB6 are required for premeiotic DNA replication and activation of the meiotic S/M checkpoint.Genes Dev. 1998 Sep 1;12(17):2698-710. doi: 10.1101/gad.12.17.2698. Genes Dev. 1998. PMID: 9732268 Free PMC article.
-
S-phase cyclin-dependent kinases promote sister chromatid cohesion in budding yeast.Mol Cell Biol. 2011 Jun;31(12):2470-83. doi: 10.1128/MCB.05323-11. Epub 2011 Apr 25. Mol Cell Biol. 2011. PMID: 21518961 Free PMC article.
-
Mechanisms involved in regulating DNA replication origins during the cell cycle and in response to DNA damage.Philos Trans R Soc Lond B Biol Sci. 2004 Jan 29;359(1441):31-8. doi: 10.1098/rstb.2003.1362. Philos Trans R Soc Lond B Biol Sci. 2004. PMID: 15065654 Free PMC article. Review.
-
Regulating DNA replication in eukarya.Cold Spring Harb Perspect Biol. 2013 Sep 1;5(9):a012930. doi: 10.1101/cshperspect.a012930. Cold Spring Harb Perspect Biol. 2013. PMID: 23838438 Free PMC article. Review.
Cited by
-
Mathematical modelling of DNA replication reveals a trade-off between coherence of origin activation and robustness against rereplication.PLoS Comput Biol. 2010 May 13;6(5):e1000783. doi: 10.1371/journal.pcbi.1000783. PLoS Comput Biol. 2010. PMID: 20485558 Free PMC article.
-
An overview of Cdk1-controlled targets and processes.Cell Div. 2010 May 13;5:11. doi: 10.1186/1747-1028-5-11. Cell Div. 2010. PMID: 20465793 Free PMC article.
-
Control of pre-replicative complex during the division cycle in Chlamydomonas reinhardtii.PLoS Genet. 2021 Apr 28;17(4):e1009471. doi: 10.1371/journal.pgen.1009471. eCollection 2021 Apr. PLoS Genet. 2021. PMID: 33909603 Free PMC article.
-
Checkpoint effects and telomere amplification during DNA re-replication in fission yeast.BMC Mol Biol. 2007 Dec 21;8:119. doi: 10.1186/1471-2199-8-119. BMC Mol Biol. 2007. PMID: 18154680 Free PMC article.
-
Method for identifying phosphorylated substrates of specific cyclin/cyclin-dependent kinase complexes.Proc Natl Acad Sci U S A. 2014 Aug 5;111(31):11323-8. doi: 10.1073/pnas.1409666111. Epub 2014 Jul 21. Proc Natl Acad Sci U S A. 2014. PMID: 25049391 Free PMC article.
References
-
- Amon, A., M. Tyers, B. Futcher and K. Nasmyth, 1993. Mechanisms that help the yeast cell cycle clock tick: G2 cyclins transcriptionally activate G2 cyclins and repress G1 cyclins. Cell 74: 993–1007. - PubMed
-
- Aparicio, O. M., D. M. Weinstein and S. P. Bell, 1997. Components and dynamics of DNA replication complexes in S. cerevisiae: redistribution of MCM proteins and Cdc45p during S phase. Cell 91: 59–69. - PubMed
-
- Archambault, V., E. J. Chang, B. J. Drapkin, F. R. Cross, B. T. Chait et al., 2004. Targeted proteomic study of the cyclin-Cdk module. Mol. Cell 14: 699–711. - PubMed
-
- Archambault, V., N. E. Buchler, G. M. Wilmes, M. D. Jacobson and F. R. Cross, 2005. a Two-faced cyclins with eyes on the targets. Cell Cycle 4: 125–130. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
