

Evidence for preferential Ig gene usage and differential TdT and exonuclease activities in human naïve and memory B cells
- PMID: 17196657
- PMCID: PMC1859862
- DOI: 10.1016/j.molimm.2006.11.020
Evidence for preferential Ig gene usage and differential TdT and exonuclease activities in human naïve and memory B cells
Abstract
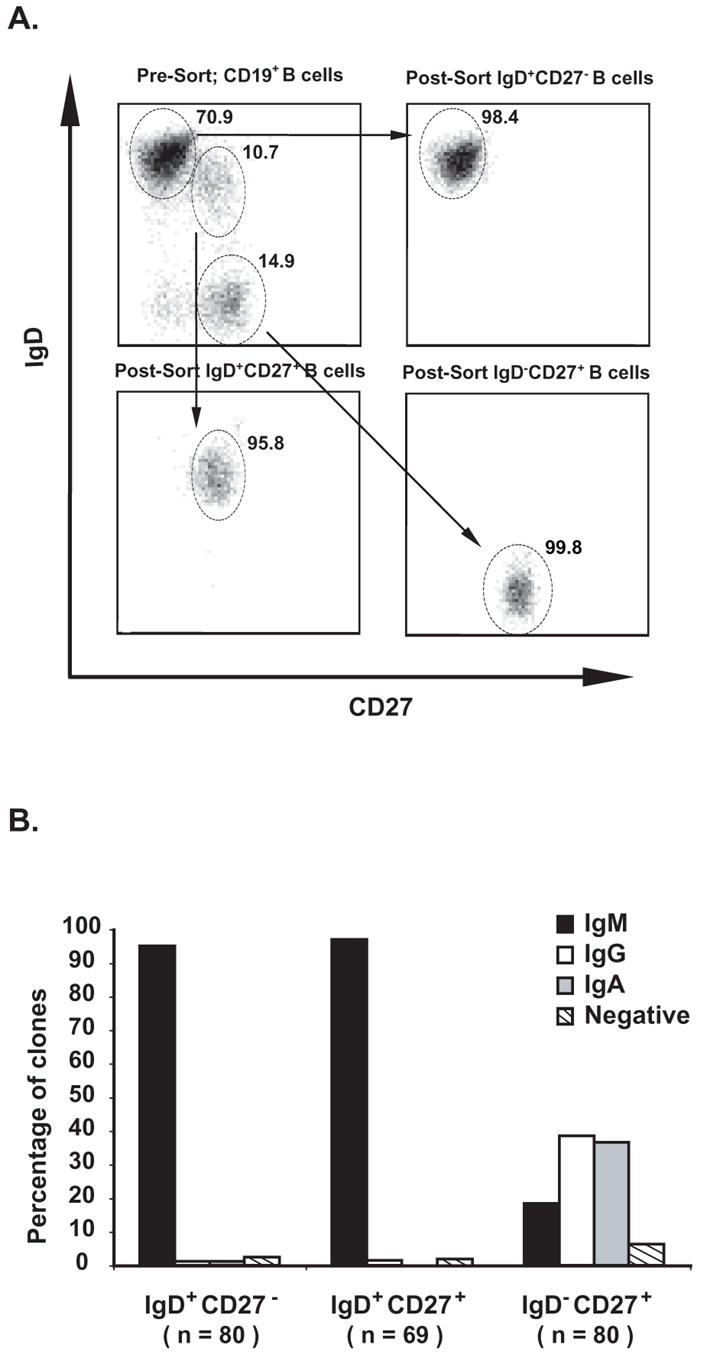
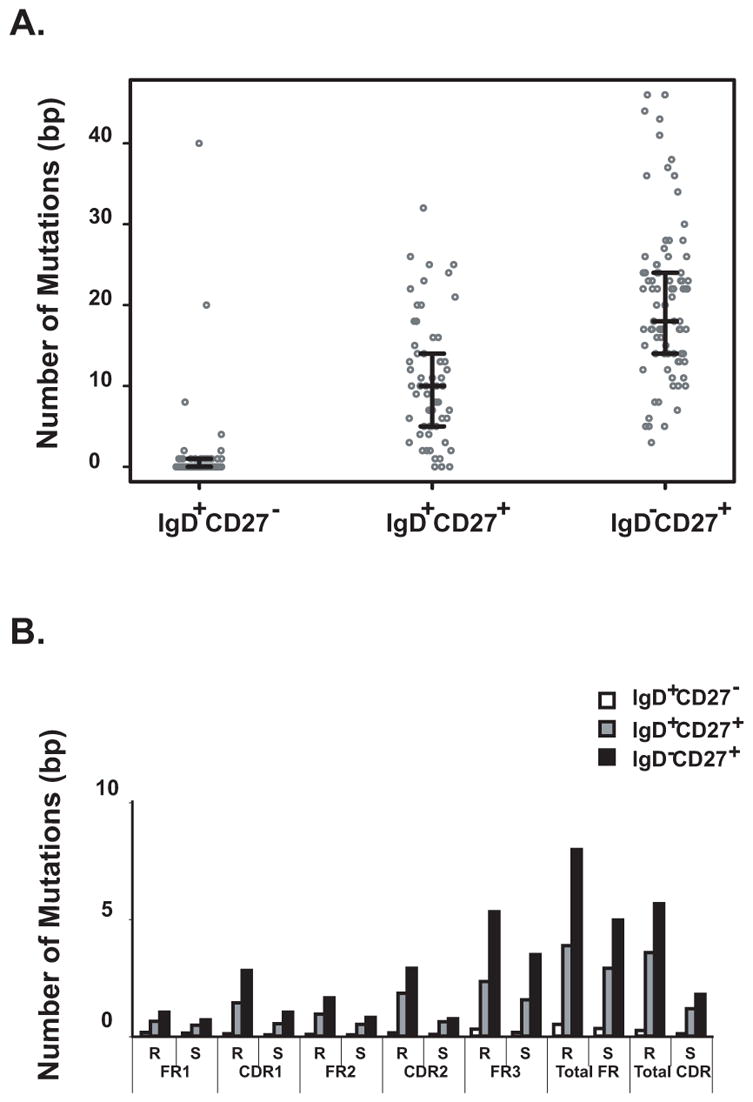
Memory B cells and the antibodies they encode are important for protective immunity against infectious pathogens. Characterization of naïve and memory B cell antibody repertoires will elucidate the molecular basis for the generation of antibody diversity in human B cells and the optimization of antibody structures that bind microbial antigens. In this study we aimed to investigate the influence of antigenic selection on the antibody genes of the two CD27+ memory B cell subsets, comparing them with the naïve repertoire in CD27- cells. We analyzed and compared the Ig heavy chain gene transcripts in three recently defined circulating naïve and memory B cell subsets (CD19+IgD+CD27- [naïve], CD19+IgD+CD27+ [un-class-switched memory] or CD19+IgD- CD27+ [class-switched memory]) at the single cell level. We found similar biased patterns of variable, diversity and joining heavy chain gene usages in all three groups of cells. CD19+IgD+CD27+ memory B cells harbored as diverse an antibody gene repertoire as CD19+IgD-CD27+ memory B cells. Interestingly, CD19+IgD+CD27+ memory B cells possessed a lower frequency of somatic mutations, a higher incidence of exonuclease activity at the 3' end of D regions, and a lower frequency of N and P nucleotide additions at both VH-D and D-JH junctions of CDR3 regions compared to CD19+IgD-CD27+ memory B cells. These data suggest distinct functional mechanisms underlying selection of this unique subset of un-class-switched memory B cells.
Figures




Similar articles
-
Immunodominance of the VH1-46 antibody gene segment in the primary repertoire of human rotavirus-specific B cells is reduced in the memory compartment through somatic mutation of nondominant clones.J Immunol. 2008 Mar 1;180(5):3279-88. doi: 10.4049/jimmunol.180.5.3279. J Immunol. 2008. PMID: 18292552
-
Abnormalities in peripheral B cell memory of patients with primary Sjögren's syndrome.Arthritis Rheum. 2004 Jun;50(6):1897-908. doi: 10.1002/art.20276. Arthritis Rheum. 2004. PMID: 15188366
-
Reduced non-switched memory B cell subsets cause imbalance in B cell repertoire in systemic sclerosis.Clin Exp Rheumatol. 2016 Sep-Oct;34 Suppl 100(5):30-36. Epub 2016 Apr 8. Clin Exp Rheumatol. 2016. PMID: 27056741
-
Memory B cells and CD27.Histol Histopathol. 2000 Apr;15(2):573-6. doi: 10.14670/HH-15.573. Histol Histopathol. 2000. PMID: 10809378 Review.
-
[Molecules involved in characteristics of naive/memory B cells].Nihon Rinsho Meneki Gakkai Kaishi. 2004 Oct;27(5):309-14. doi: 10.2177/jsci.27.309. Nihon Rinsho Meneki Gakkai Kaishi. 2004. PMID: 15559319 Review. Japanese.
Cited by
-
Low frequency of broadly neutralizing HIV antibodies during chronic infection even in quaternary epitope targeting antibodies containing large numbers of somatic mutations.Mol Immunol. 2016 Feb;70:94-103. doi: 10.1016/j.molimm.2015.12.002. Epub 2015 Dec 31. Mol Immunol. 2016. PMID: 26748387 Free PMC article.
-
Ontogeny-based immunogens for the induction of V2-directed HIV broadly neutralizing antibodies.Immunol Rev. 2017 Jan;275(1):217-229. doi: 10.1111/imr.12501. Immunol Rev. 2017. PMID: 28133797 Free PMC article. Review.
-
Loss of B Cells in Patients with Heterozygous Mutations in IKAROS.N Engl J Med. 2016 Mar 17;374(11):1032-1043. doi: 10.1056/NEJMoa1512234. N Engl J Med. 2016. PMID: 26981933 Free PMC article.
-
Multiple Antibody Lineages in One Donor Target the Glycan-V3 Supersite of the HIV-1 Envelope Glycoprotein and Display a Preference for Quaternary Binding.J Virol. 2016 Nov 14;90(23):10574-10586. doi: 10.1128/JVI.01012-16. Print 2016 Dec 1. J Virol. 2016. PMID: 27654288 Free PMC article.
-
High-throughput immunoglobulin repertoire analysis distinguishes between human IgM memory and switched memory B-cell populations.Blood. 2010 Aug 19;116(7):1070-8. doi: 10.1182/blood-2010-03-275859. Epub 2010 May 10. Blood. 2010. PMID: 20457872 Free PMC article.
References
-
- Agematsu K. Memory B cells and CD27. Histol Histopathol. 2000;15:573–6. - PubMed
-
- Agematsu K, Hokibara S, Nagumo H, Komiyama A. CD27: a memory B-cell marker. Immunol Today. 2000;21:204–6. - PubMed
-
- Andria ML, Levy S, Benjamini E. Diverse VH and VL genes are used to produce antibodies against a defined protein epitope. J Immunol. 1990;144:2614–9. - PubMed
-
- Berek C, Milstein C. Mutation drift and repertoire shift in the maturation of the immune response. Immunol Rev. 1987;96:23–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
