

Identification and functional characterization of the hepatic stellate cell CD38 cell surface molecule
- PMID: 17200192
- PMCID: PMC1762705
- DOI: 10.2353/ajpath.2007.051212
Identification and functional characterization of the hepatic stellate cell CD38 cell surface molecule
Abstract
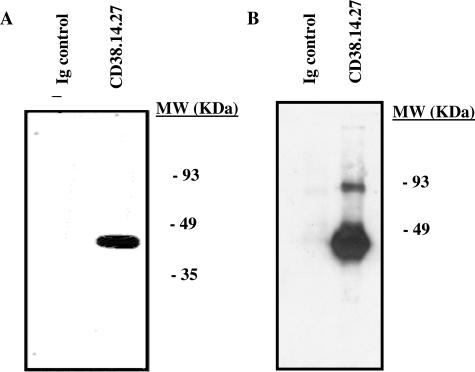
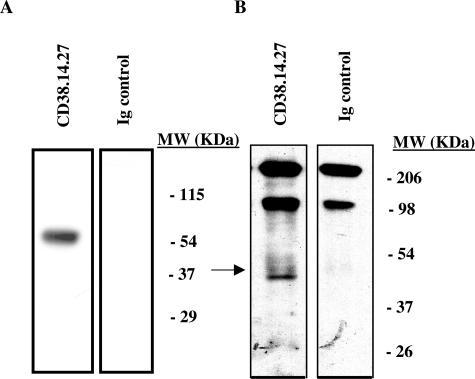
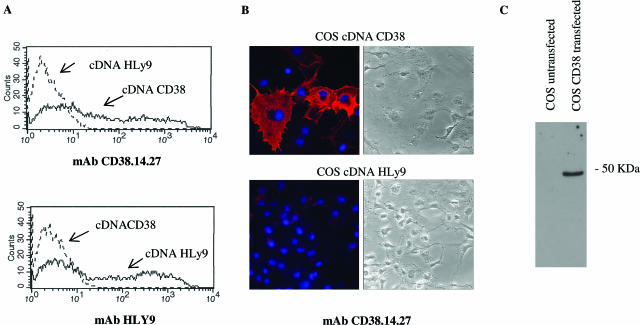
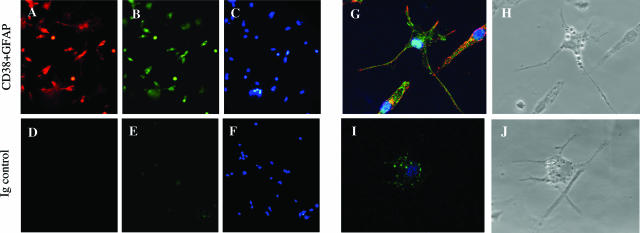
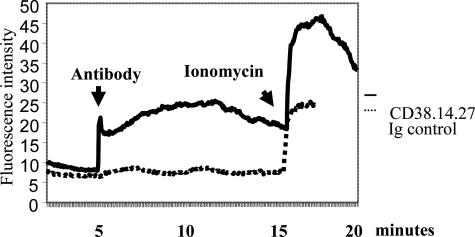
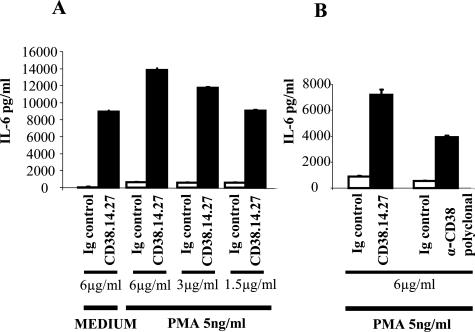
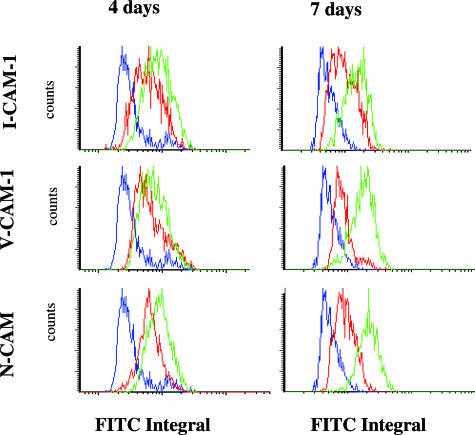
The activation of hepatic stellate cells (HSCs) is a critical event in hepatic fibrosis, because these cells are the main producers of extracellular matrix proteins in the liver and contribute to the modulation of inflammatory responses via the secretion of several cytokines and the expression of adhesion molecules. The goal of the present study was to characterize cell surface proteins that regulate HSC activation. To this end, a panel of monoclonal antibodies (mAbs) was generated. mAb 14.27 recognized a protein of 45 kd that was highly expressed on HSCs. Affinity purification of this protein followed by sequencing revealed that protein to be CD38. We subsequently demonstrated that CD38 was constitutively expressed by HSCs and that its expression increased after in vitro and in vivo activation. mAb 14.27 induced an increase in cytosolic Ca2+ levels in HSCs, showing that it functions as an agonistic antibody. Moreover, the effects mediated by the CD38 mAb included induction of the proinflammatory cytokine interleukin-6 and up-regulation of the adhesion molecules intercellular adhesion molecule-1, vascular cell adhesion molecule-1, and neural cell adhesion molecule. Collectively, our data suggest that CD38 can act as a regulator of HSC activation and effector functions.
Figures

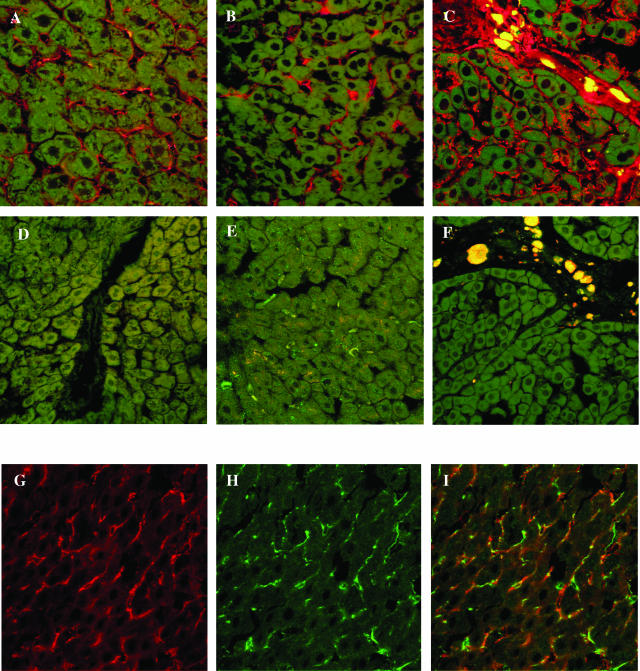
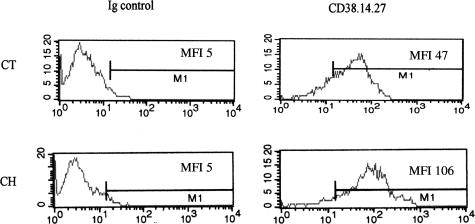
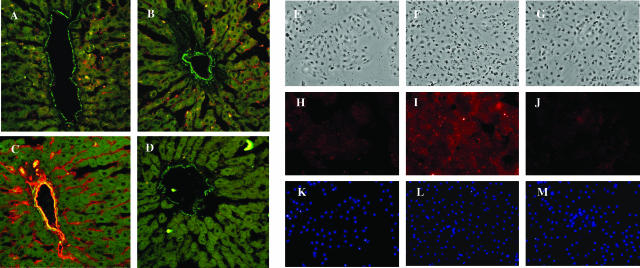
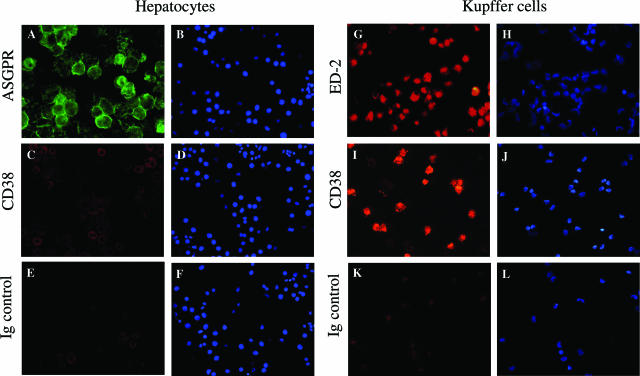
References
-
- Geerts A. History, heterogeneity, developmental biology, and functions of quiescent hepatic stellate cells. Semin Liver Dis. 2001;21:311–335. - PubMed
-
- Friedman SL. Molecular regulation of hepatic fibrosis, an integrated cellular response to tissue injury. J Biol Chem. 2000;275:2247–2250. - PubMed
-
- Albanis E, Friedman SL. Hepatic fibrosis: pathogenesis and principles of therapy. Clin Liver Dis. 2001;5:315–334, v-vi. - PubMed
-
- Friedman SL. Seminars in medicine of the Beth Israel Hospital, Boston. The cellular basis of hepatic fibrosis: mechanisms and treatment strategies. N Engl J Med. 1993;328:1828–1835. - PubMed
-
- Bataller R, North KE, Brenner DA. Genetic polymorphisms and the progression of liver fibrosis: a critical appraisal. Hepatology. 2003;37:493–503. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
