

Serum response factor binding sites differ in three human cell types
- PMID: 17200232
- PMCID: PMC1781345
- DOI: 10.1101/gr.5875007
Serum response factor binding sites differ in three human cell types
Abstract
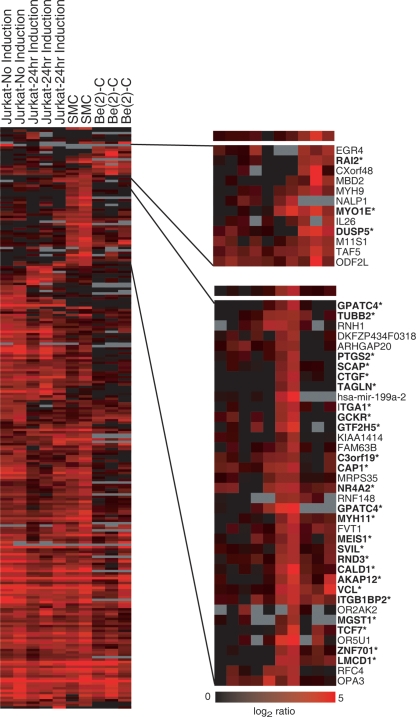
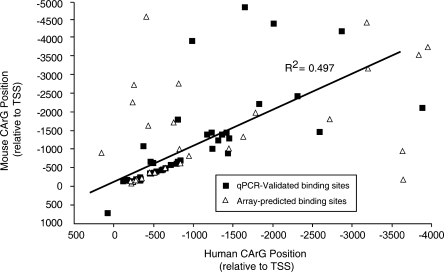
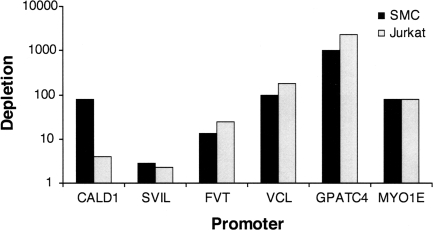
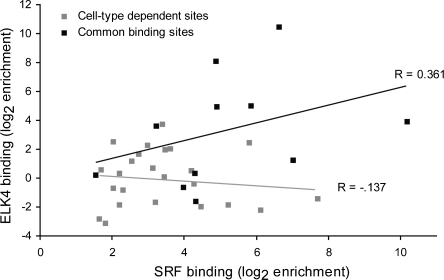
The serum response factor (SRF) is essential for embryonic development and maintenance of muscle cells and neurons. The mechanism by which this factor controls these divergent pathways is unclear. Here we present a genome-wide view of occupancy of SRF at its binding sites with a focus on those that vary with cell type. We used chromatin immunoprecipitation (ChIP) in combination with human promoter microarrays to identify 216 putative SRF binding sites in the human genome. We performed independent quantitative PCR validation at over half of these sites that resulted in 146 sites we assert to be true binding sites at over 90% confidence. Nearly half of the sites are bound by SRF in only one of the three cell types we tested, providing strong evidence for the diverse roles for SRF in different cell types. We also explore possible mechanisms controlling differential binding of SRF in these cell types by assaying cofactor binding, DNA methylation, histone methylation, and histone acetylation at a subset of sites bound preferentially in smooth muscle cells. Although we did not see a strong correlation between SRF binding and epigenetics modifications, at these sites, we propose that SRF cofactors may play an important role in determining cell-dependent SRF binding sites. ELK4 (previously known as SAP-1 [SRF-associated protein-1]) is ubiquitously expressed. Therefore, we expected it to occupy sites where SRF binding is common in all cell types. Indeed, 90% of SRF sites also bound by ELK4 were common to all three cell types. Together, our data provide a more complete understanding of the regulatory network controlled by SRF.
Figures
References
-
- Alberti S., Krause S.M., Kretz O., Philippar U., Lemberger T., Casanova E., Wiebel F.F., Schwarz H., Frotscher M., Schutz G., Krause S.M., Kretz O., Philippar U., Lemberger T., Casanova E., Wiebel F.F., Schwarz H., Frotscher M., Schutz G., Kretz O., Philippar U., Lemberger T., Casanova E., Wiebel F.F., Schwarz H., Frotscher M., Schutz G., Philippar U., Lemberger T., Casanova E., Wiebel F.F., Schwarz H., Frotscher M., Schutz G., Lemberger T., Casanova E., Wiebel F.F., Schwarz H., Frotscher M., Schutz G., Casanova E., Wiebel F.F., Schwarz H., Frotscher M., Schutz G., Wiebel F.F., Schwarz H., Frotscher M., Schutz G., Schwarz H., Frotscher M., Schutz G., Frotscher M., Schutz G., Schutz G., et al. Neuronal migration in the murine rostral migratory stream requires serum response factor. Proc. Natl. Acad. Sci. 2005;102:6148–6153. - PMC - PubMed
-
- Arsenian S., Weinhold B., Oelgeschlager M., Ruther U., Nordheim A., Weinhold B., Oelgeschlager M., Ruther U., Nordheim A., Oelgeschlager M., Ruther U., Nordheim A., Ruther U., Nordheim A., Nordheim A. Serum response factor is essential for mesoderm formation during mouse embryogenesis. EMBO J. 1998;17:6289–6299. - PMC - PubMed
-
- Balza R.O., Jr., Misra R.P., Misra R.P. Role of the serum response factor in regulating contractile apparatus gene expression and sarcomeric integrity in cardiomyocytes. J. Biol. Chem. 2006;281:6498–6510. - PubMed
-
- Belaguli N.S., Schildmeyer L.A., Schwartz R.J., Schildmeyer L.A., Schwartz R.J., Schwartz R.J. Organization and myogenic restricted expression of the murine serum response factor gene. A role for autoregulation. J. Biol. Chem. 1997;272:18222–18231. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous