

Spatial targeting of type II protein kinase A to filopodia mediates the regulation of growth cone guidance by cAMP
- PMID: 17200417
- PMCID: PMC2063631
- DOI: 10.1083/jcb.200607128
Spatial targeting of type II protein kinase A to filopodia mediates the regulation of growth cone guidance by cAMP
Abstract
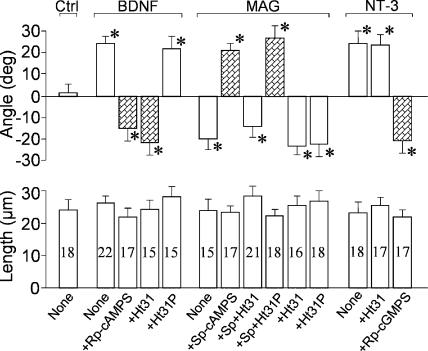
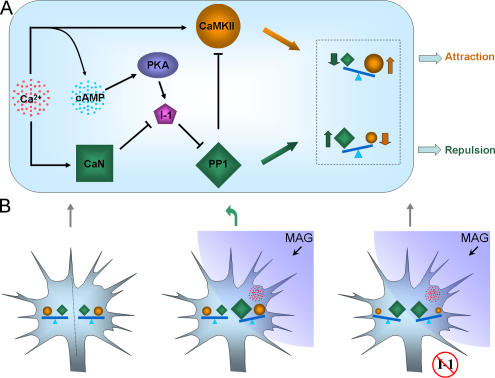
The second messenger cyclic adenosine monophosphate (cAMP) plays a pivotal role in axonal growth and guidance, but its downstream mechanisms remain elusive. In this study, we report that type II protein kinase A (PKA) is highly enriched in growth cone filopodia, and this spatial localization enables the coupling of cAMP signaling to its specific effectors to regulate guidance responses. Disrupting the localization of PKA to filopodia impairs cAMP-mediated growth cone attraction and prevents the switching of repulsive responses to attraction by elevated cAMP. Our data further show that PKA targets protein phosphatase-1 (PP1) through the phosphorylation of a regulatory protein inhibitor-1 (I-1) to promote growth cone attraction. Finally, we find that I-1 and PP1 mediate growth cone repulsion induced by myelin-associated glycoprotein. These findings demonstrate that the spatial localization of type II PKA to growth cone filopodia plays an important role in the regulation of growth cone motility and guidance by cAMP.
Figures







References
-
- Aubert, I., J.L. Ridet, and F.H. Gage. 1995. Regeneration in the adult mammalian CNS: guided by development. Curr. Opin. Neurobiol. 5:625–635. - PubMed
-
- Bahr, M., and F. Bonhoeffer. 1994. Perspectives on axonal regeneration in the mammalian CNS. Trends Neurosci. 17:473–479. - PubMed
-
- Banker, G.A., and W.M. Cowan. 1977. Rat hippocampal neurons in dispersed cell culture. Brain Res. 126:397–425. - PubMed
-
- Bauman, A.L., A.S. Goehring, and J.D. Scott. 2004. Orchestration of synaptic plasticity through AKAP signaling complexes. Neuropharmacology. 46:299–310. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
