

Quaternary structure and biochemical properties of mycobacterial RNase E/G
- PMID: 17201693
- PMCID: PMC1828891
- DOI: 10.1042/BJ20061530
Quaternary structure and biochemical properties of mycobacterial RNase E/G
Abstract
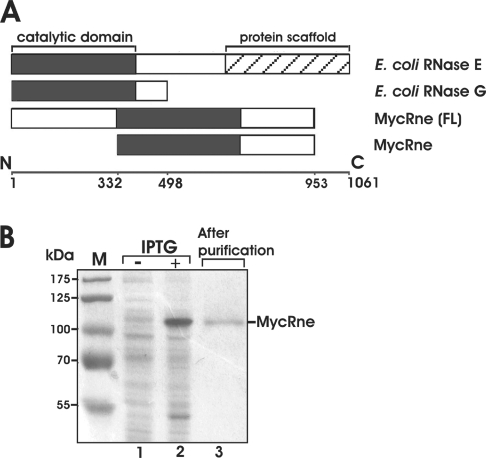
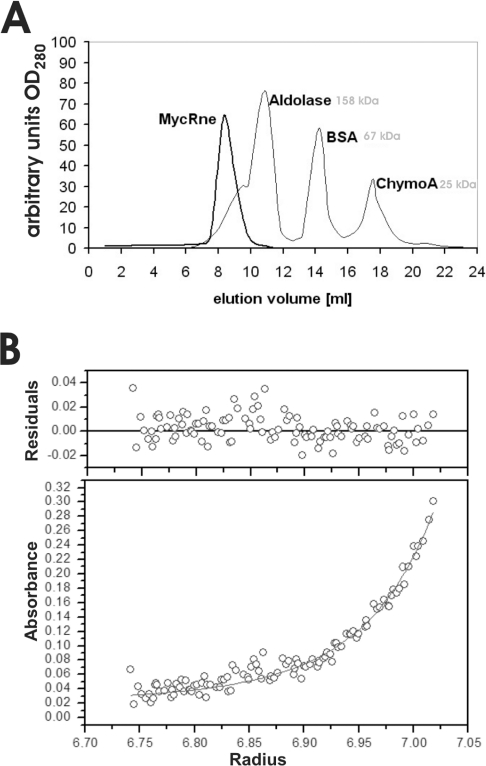
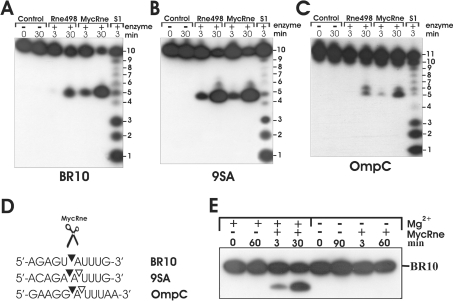
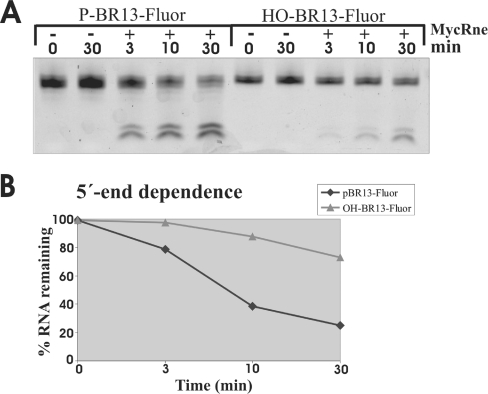
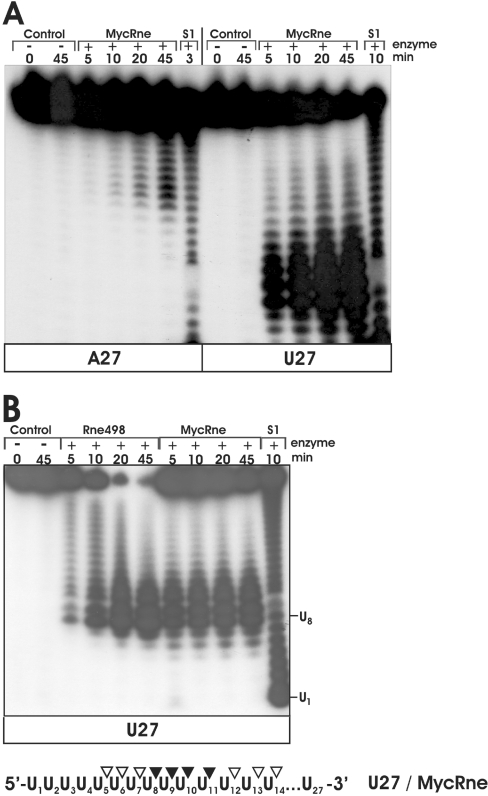
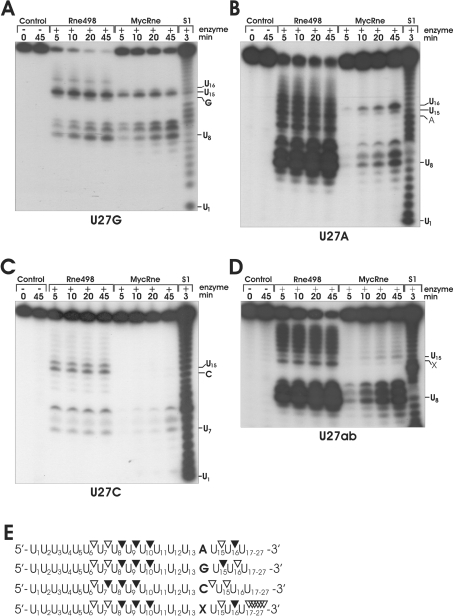
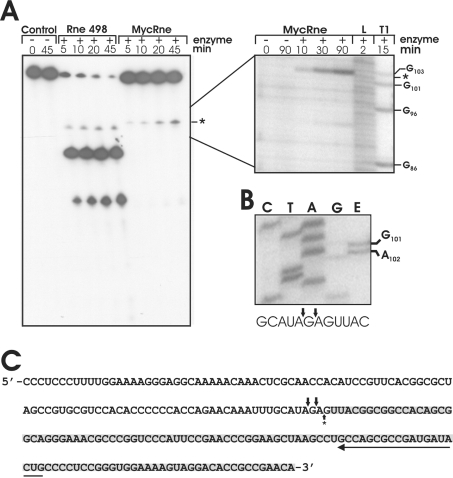
The RNase E/G family of endoribonucleases plays the central role in numerous post-transcriptional mechanisms in Escherichia coli and, presumably, in other bacteria, including human pathogens. To learn more about specific properties of RNase E/G homologues from pathogenic Gram-positive bacteria, a polypeptide comprising the catalytic domain of Mycobacterium tuberculosis RNase E/G (MycRne) was purified and characterized in vitro. In the present study, we show that affinity-purified MycRne has a propensity to form dimers and tetramers in solution and possesses an endoribonucleolytic activity, which is dependent on the 5'-phosphorylation status of RNA. Our data also indicate that the cleavage specificities of the M. tuberculosis RNase E/G homologue and its E. coli counterpart are only moderately overlapping, and reveal a number of sequence determinants within MycRne cleavage sites that differentially affect the efficiency of cleavage. Finally, we demonstrate that, similar to E. coli RNase E, MycRne is able to cleave in an intercistronic region of the putative 9S precursor of 5S rRNA, thus suggesting a common function for RNase E/G homologues in rRNA processing.
Figures
References
-
- Cohen S. N., McDowall K. J. RNase E: still a wonderfully mysterious enzyme. Mol. Microbiol. 1997;23:1099–1106. - PubMed
-
- Coburn G. A., Mackie G. A. Degradation of mRNA in Escherichia coli: an old problem with some new twists. Prog. Nucleic Acids Res. Mol. Biol. 1999;62:55–108. - PubMed
-
- Régnier P., Arraiano C. M. Degradation of mRNA in bacteria: emergence of ubiquitous features. BioEssays. 2000;22:235–244. - PubMed
-
- Deana A., Belasco J. G. The function of RNase G in Escherichia coli is constrained by its amino and carboxyl termini. Mol. Microbiol. 2004;51:1205–1217. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources