

The glucose transporter 4-regulating protein TUG is essential for highly insulin-responsive glucose uptake in 3T3-L1 adipocytes
- PMID: 17202135
- PMCID: PMC2243252
- DOI: 10.1074/jbc.M610824200
The glucose transporter 4-regulating protein TUG is essential for highly insulin-responsive glucose uptake in 3T3-L1 adipocytes
Abstract
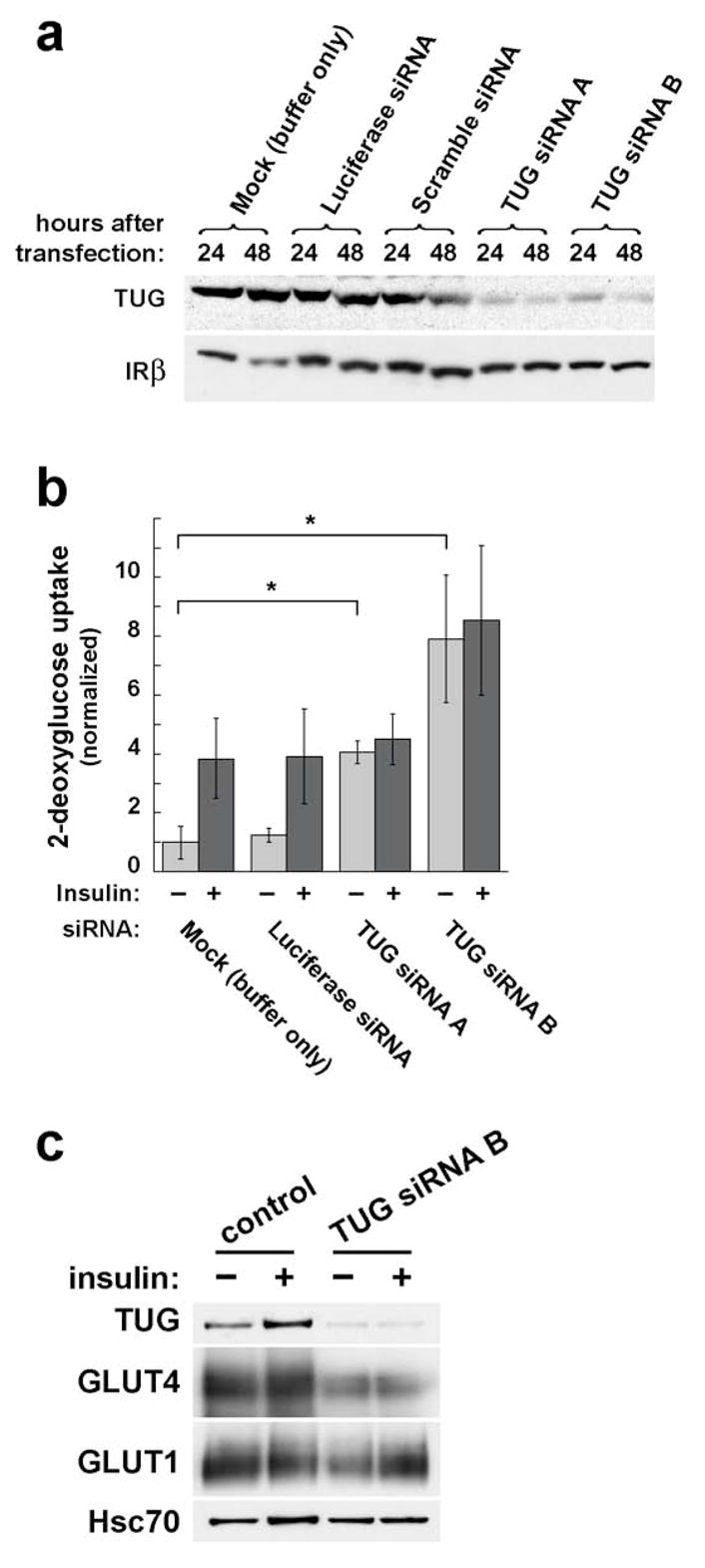
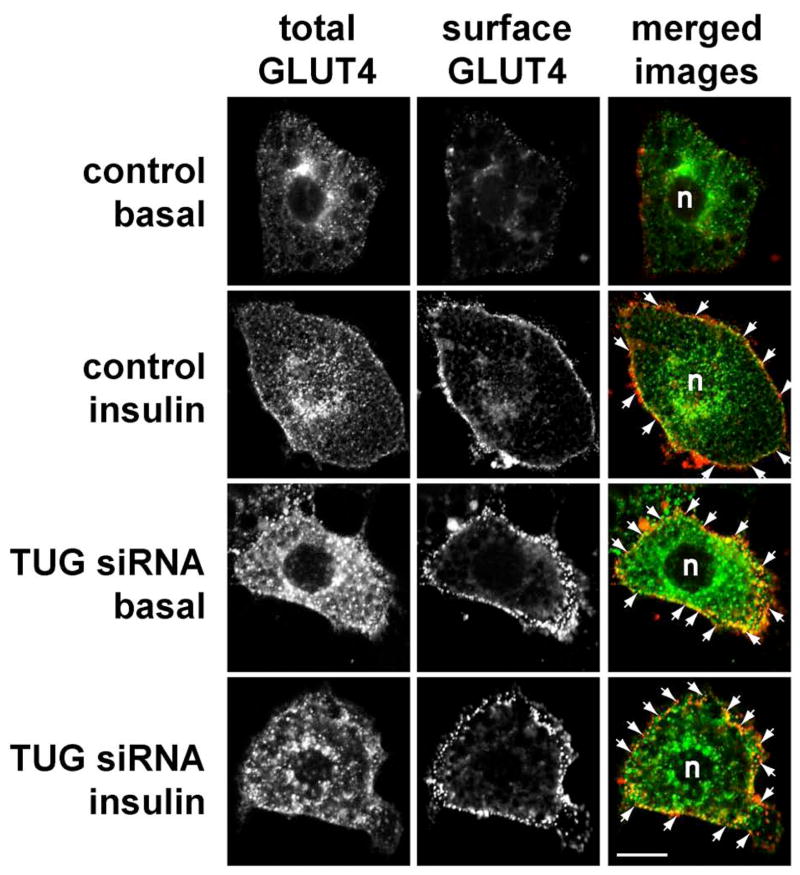
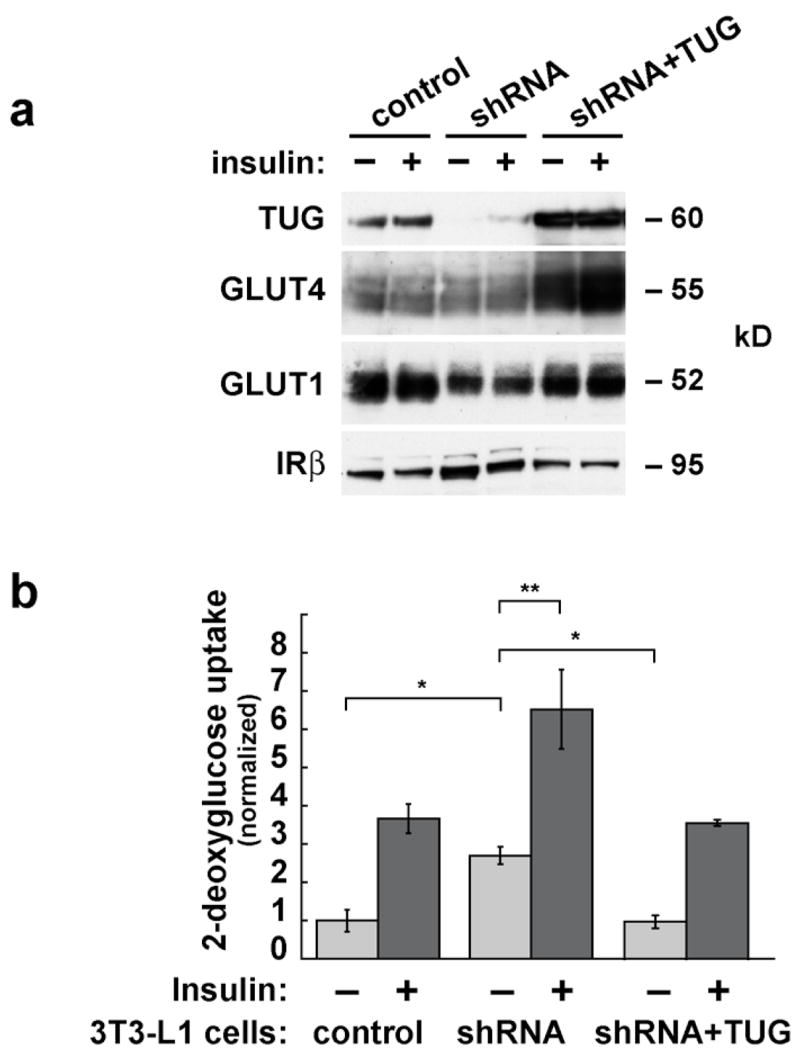
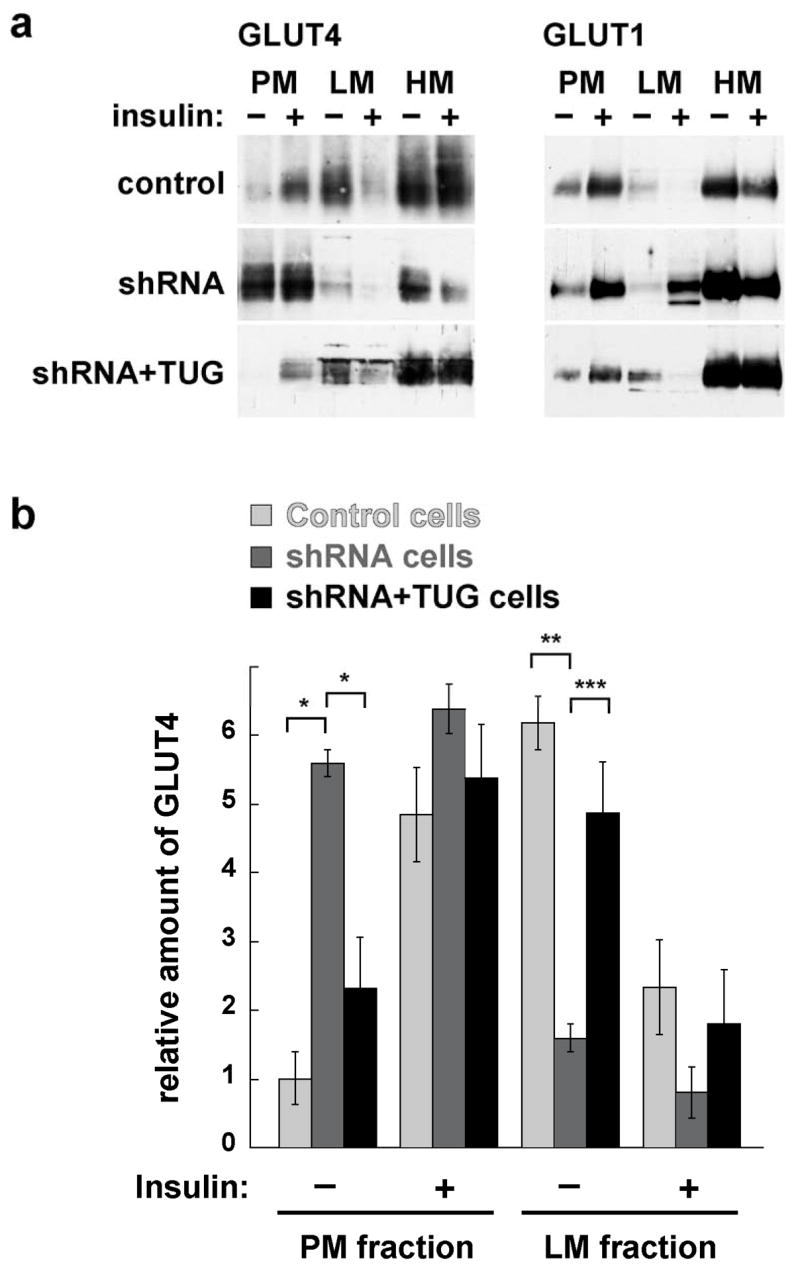
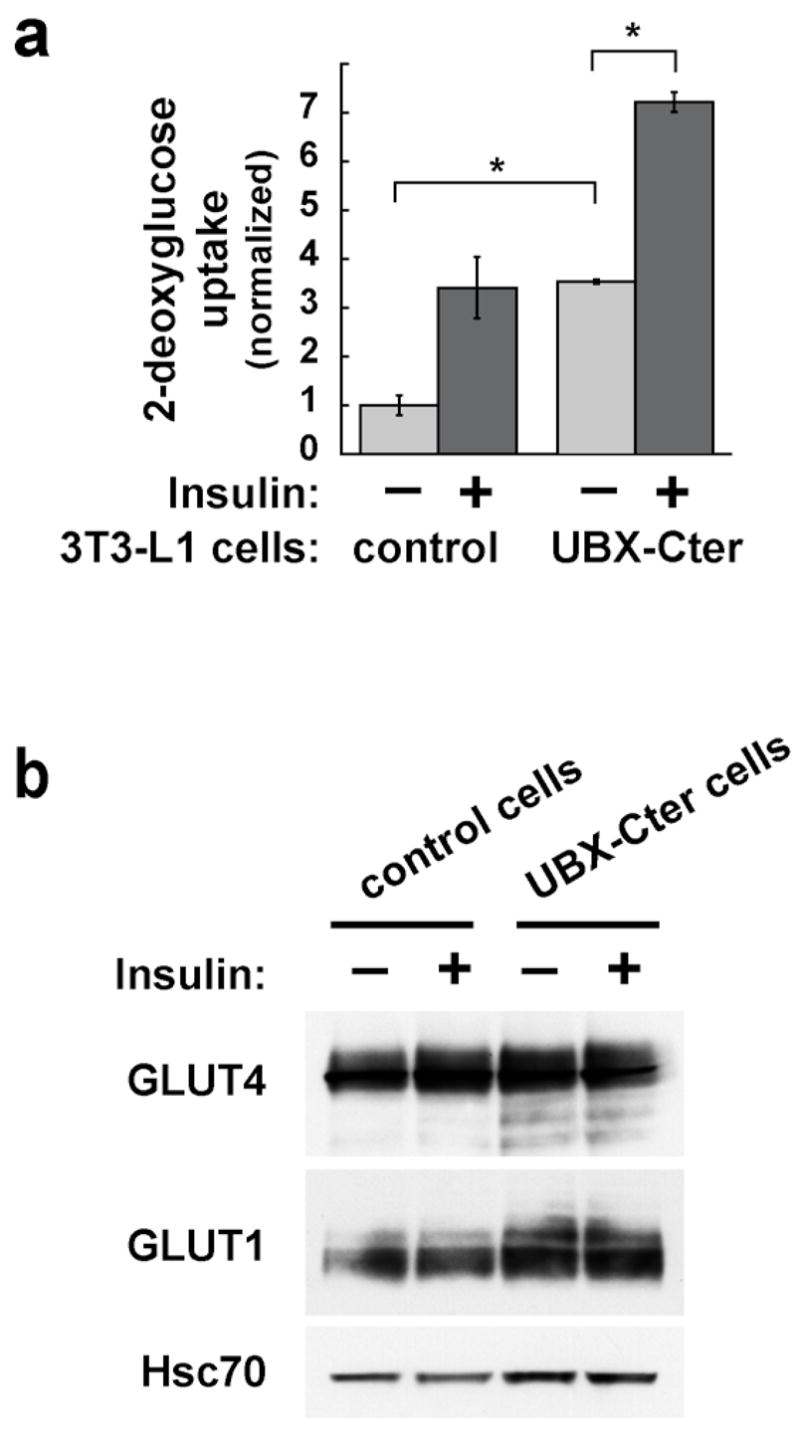
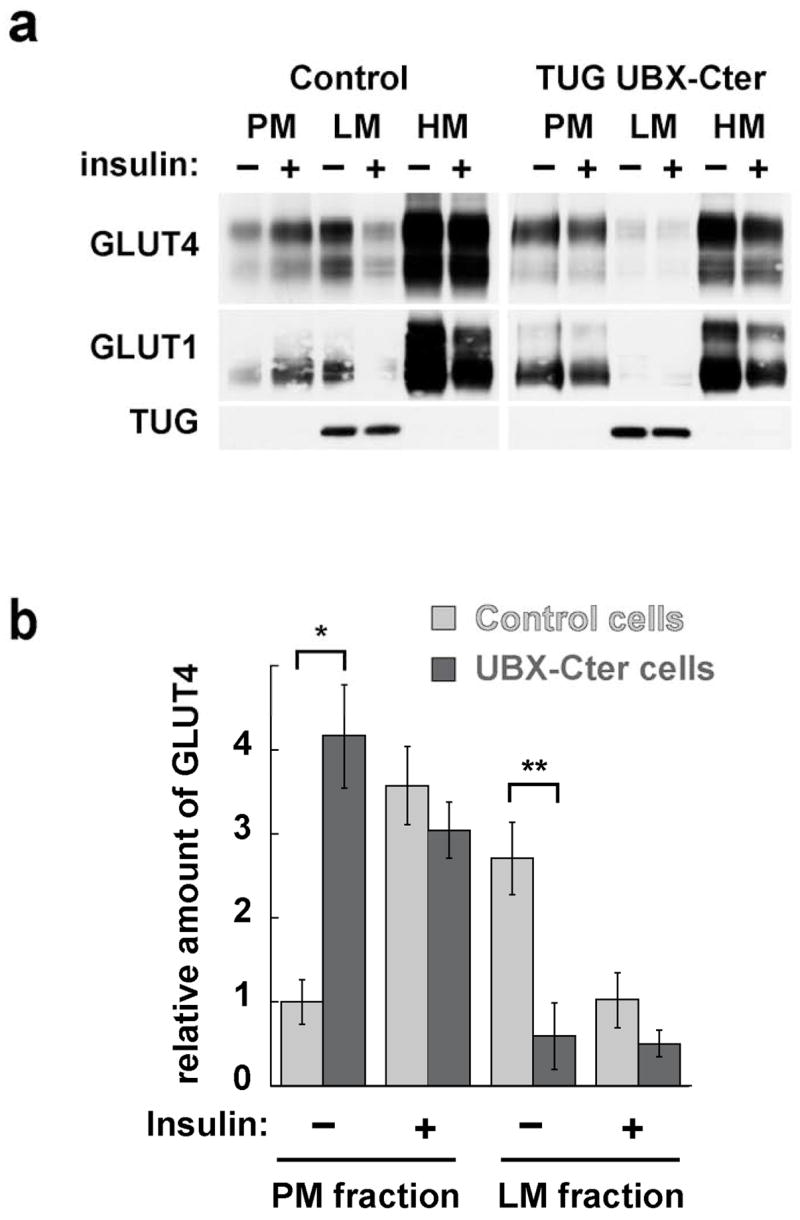
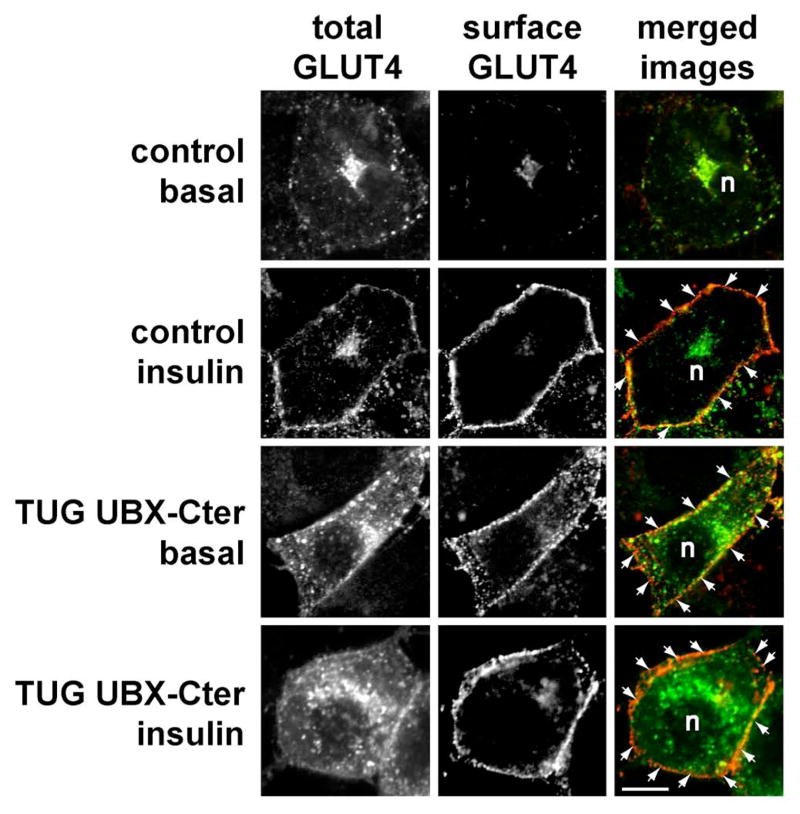
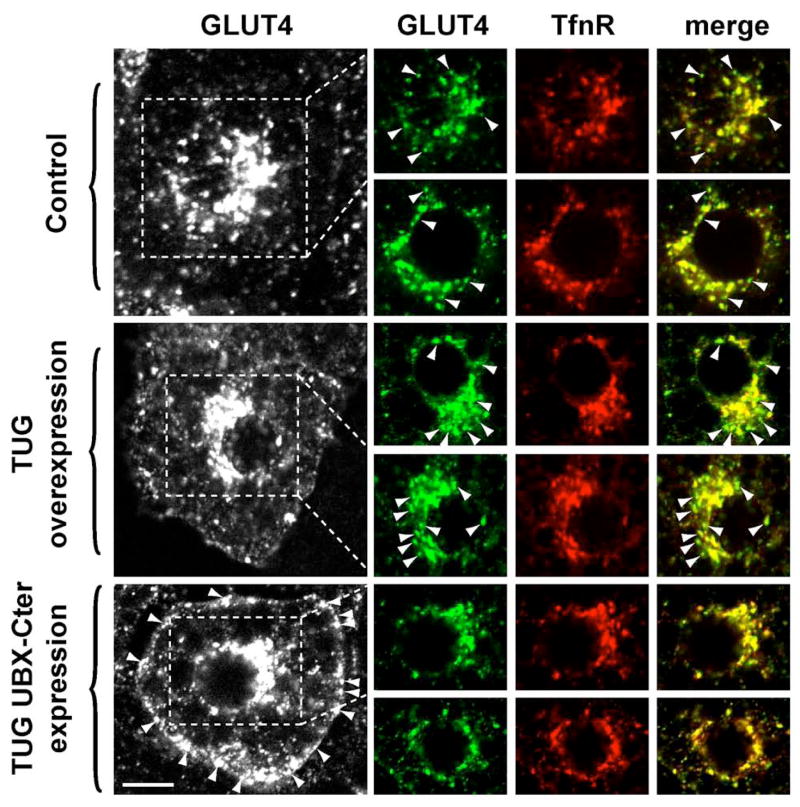
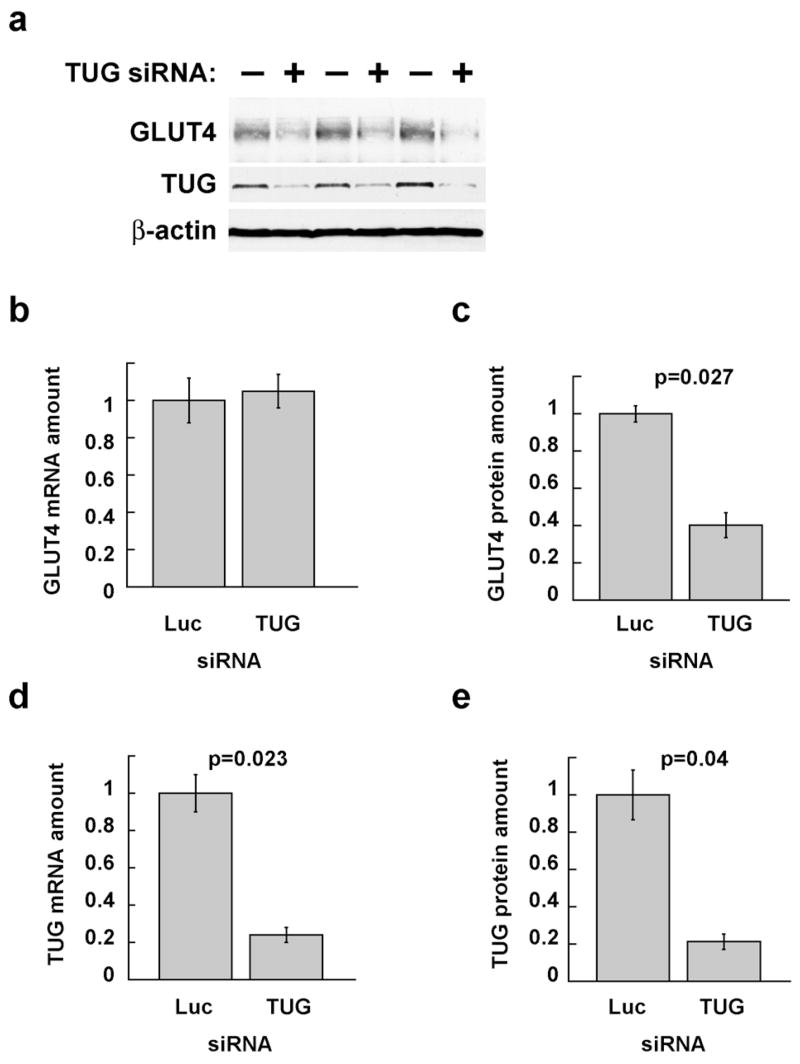
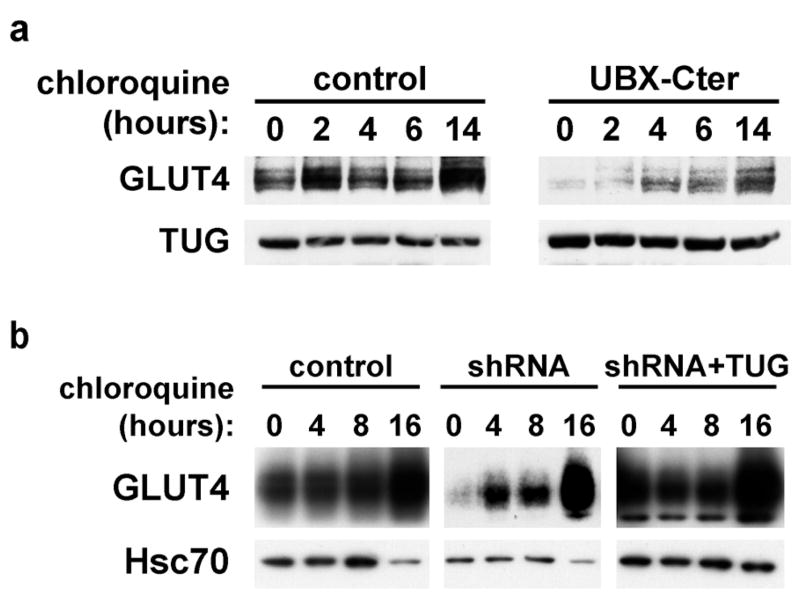
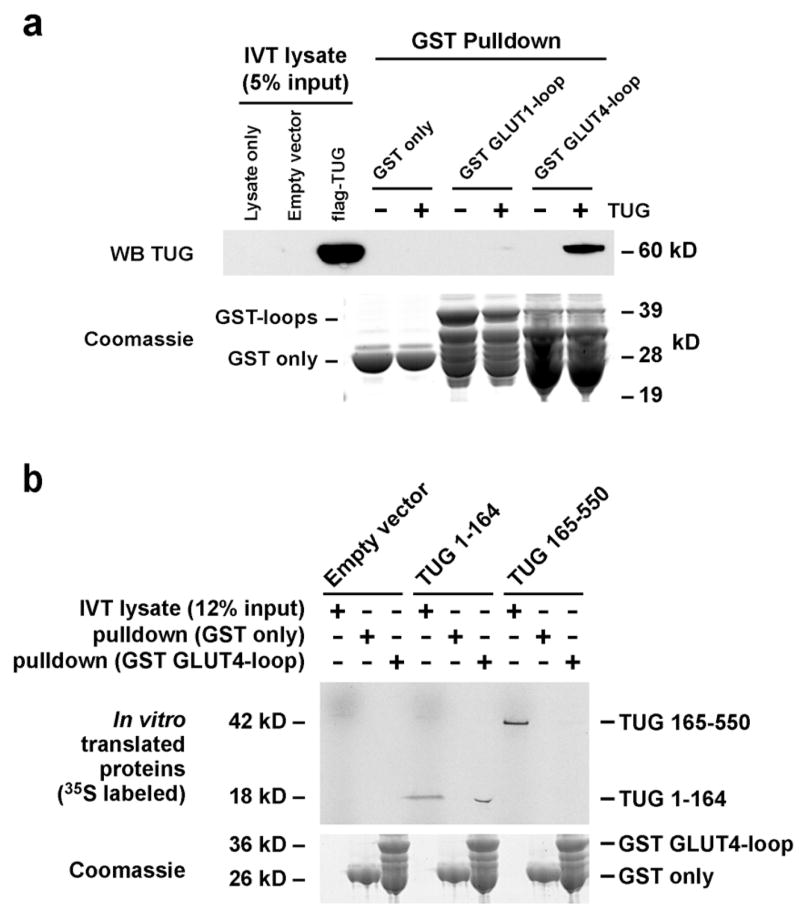
Insulin stimulates glucose uptake in fat and muscle by redistributing GLUT4 glucose transporters from intracellular membranes to the cell surface. We previously proposed that, in 3T3-L1 adipocytes, TUG retains GLUT4 within unstimulated cells and insulin mobilizes this retained GLUT4 by stimulating its dissociation from TUG. Yet the relative importance of this action in the overall control of glucose uptake remains uncertain. Here we report that transient, small interfering RNA-mediated depletion of TUG causes GLUT4 translocation and enhances glucose uptake in unstimulated 3T3-L1 adipocytes, similar to insulin. Stable TUG depletion or expression of a dominant negative fragment likewise stimulates GLUT4 redistribution and glucose uptake, and insulin causes a 2-fold further increase. Microscopy shows that TUG governs the accumulation of GLUT4 in perinuclear membranes distinct from endosomes and indicates that it is this pool of GLUT4 that is mobilized by TUG disruption. Interestingly, in addition to translocating GLUT4 and enhancing glucose uptake, TUG disruption appears to accelerate the degradation of GLUT4 in lysosomes. Finally, we find that TUG binds directly and specifically to a large intracellular loop in GLUT4. Together, these findings demonstrate that TUG is required to retain GLUT4 intracellularly in 3T3-L1 adipocytes in the absence of insulin and further implicate the insulin-stimulated dissociation of TUG and GLUT4 as an important action by which insulin stimulates glucose uptake.
Figures
Similar articles
-
Endoproteolytic cleavage of TUG protein regulates GLUT4 glucose transporter translocation.J Biol Chem. 2012 Jul 6;287(28):23932-47. doi: 10.1074/jbc.M112.339457. Epub 2012 May 18. J Biol Chem. 2012. PMID: 22610098 Free PMC article.
-
Acetylation of TUG protein promotes the accumulation of GLUT4 glucose transporters in an insulin-responsive intracellular compartment.J Biol Chem. 2015 Feb 13;290(7):4447-63. doi: 10.1074/jbc.M114.603977. Epub 2015 Jan 5. J Biol Chem. 2015. PMID: 25561724 Free PMC article.
-
Functional cloning of TUG as a regulator of GLUT4 glucose transporter trafficking.Nature. 2003 Oct 16;425(6959):727-33. doi: 10.1038/nature01989. Nature. 2003. PMID: 14562105
-
Biogenesis and regulation of insulin-responsive vesicles containing GLUT4.Curr Opin Cell Biol. 2010 Aug;22(4):506-12. doi: 10.1016/j.ceb.2010.03.012. Epub 2010 Apr 21. Curr Opin Cell Biol. 2010. PMID: 20417083 Free PMC article. Review.
-
The molecular basis of insulin-stimulated glucose uptake: signalling, trafficking and potential drug targets.J Endocrinol. 2009 Oct;203(1):1-18. doi: 10.1677/JOE-09-0037. Epub 2009 Apr 23. J Endocrinol. 2009. PMID: 19389739 Review.
Cited by
-
Coordinated Regulation of Vasopressin Inactivation and Glucose Uptake by Action of TUG Protein in Muscle.J Biol Chem. 2015 Jun 5;290(23):14454-61. doi: 10.1074/jbc.C115.639203. Epub 2015 May 5. J Biol Chem. 2015. PMID: 25944897 Free PMC article.
-
Regulation of glucose transport by insulin: traffic control of GLUT4.Nat Rev Mol Cell Biol. 2012 May 23;13(6):383-96. doi: 10.1038/nrm3351. Nat Rev Mol Cell Biol. 2012. PMID: 22617471 Review.
-
Optogenetic activation reveals distinct roles of PIP3 and Akt in adipocyte insulin action.J Cell Sci. 2016 May 15;129(10):2085-95. doi: 10.1242/jcs.174805. Epub 2016 Apr 13. J Cell Sci. 2016. PMID: 27076519 Free PMC article.
-
UBXN9 governs GLUT4-mediated spatial confinement of RIG-I-like receptors and signaling.Res Sq [Preprint]. 2024 Jun 4:rs.3.rs-3373803. doi: 10.21203/rs.3.rs-3373803/v1. Res Sq. 2024. Update in: Nat Immunol. 2024 Dec;25(12):2234-2246. doi: 10.1038/s41590-024-02004-7. PMID: 38883790 Free PMC article. Updated. Preprint.
-
GLUT4 On the move.Biochem J. 2022 Feb 11;479(3):445-462. doi: 10.1042/BCJ20210073. Biochem J. 2022. PMID: 35147164 Free PMC article.
References
-
- Bryant NJ, Govers R, James DE. Nat Rev Mol Cell Biol. 2002;3(4):267–277. - PubMed
-
- Watson RT, Kanzaki M, Pessin JE. Endocr Rev. 2004;25(2):177–204. - PubMed
-
- Abel ED, Shepherd PR, Kahn BB. Glucose transporters and pathophysiologic states. In: Le Roith D, Taylor SI, Olefsky JM, editors. Diabetes mellitus: a fundamental and clinical text. 3. Lippincott-Raven; Philadelphia: 2003.
-
- Maianu L, Keller SR, Garvey WT. J Clin Endocrinol Metab. 2001;86(11):5450–5456. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
