

A structured viroid RNA serves as a substrate for dicer-like cleavage to produce biologically active small RNAs but is resistant to RNA-induced silencing complex-mediated degradation
- PMID: 17202210
- PMCID: PMC1865973
- DOI: 10.1128/JVI.02339-06
A structured viroid RNA serves as a substrate for dicer-like cleavage to produce biologically active small RNAs but is resistant to RNA-induced silencing complex-mediated degradation
Abstract
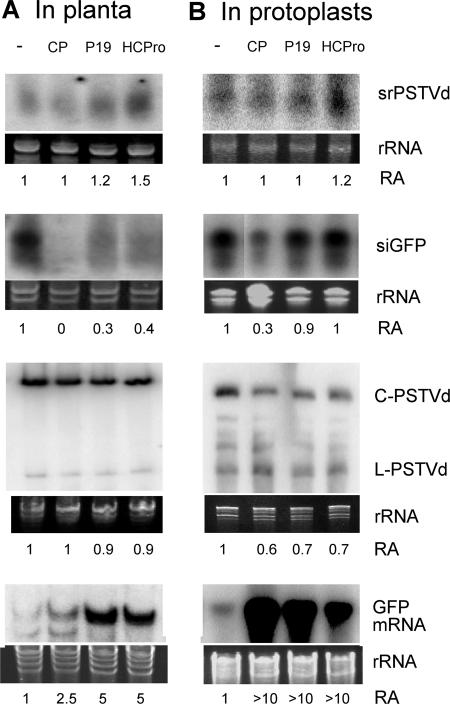
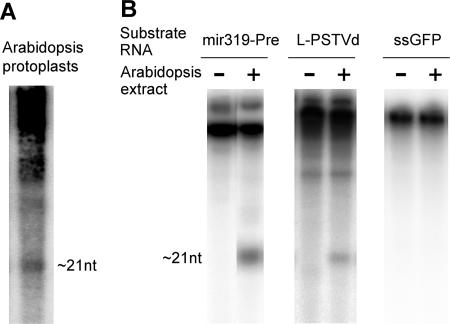
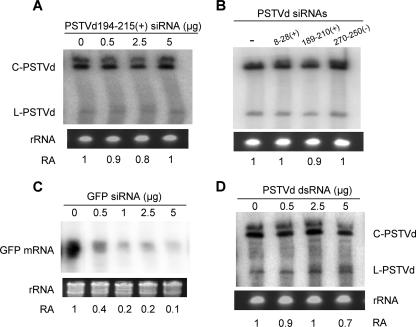
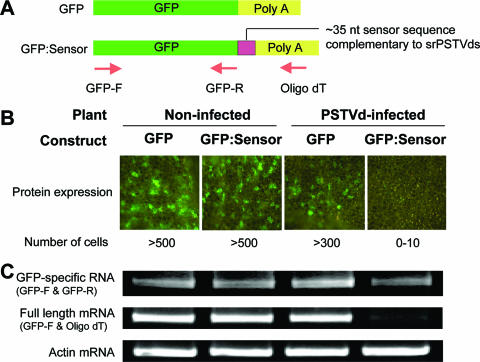
RNA silencing is a potent means of antiviral defense in plants and animals. A hallmark of this defense response is the production of 21- to 24-nucleotide viral small RNAs via mechanisms that remain to be fully understood. Many viruses encode suppressors of RNA silencing, and some viral RNAs function directly as silencing suppressors as counterdefense. The occurrence of viroid-specific small RNAs in infected plants suggests that viroids can trigger RNA silencing in a host, raising the question of how these noncoding and unencapsidated RNAs survive cellular RNA-silencing systems. We address this question by characterizing the production of small RNAs of Potato spindle tuber viroid (srPSTVds) and investigating how PSTVd responds to RNA silencing. Our molecular and biochemical studies provide evidence that srPSTVds were derived mostly from the secondary structure of viroid RNAs. Replication of PSTVd was resistant to RNA silencing, although the srPSTVds were biologically active in guiding RNA-induced silencing complex (RISC)-mediated cleavage, as shown with a sensor system. Further analyses showed that without possessing or triggering silencing suppressor activities, the PSTVd secondary structure played a critical role in resistance to RISC-mediated cleavage. These findings support the hypothesis that some infectious RNAs may have evolved specific secondary structures as an effective means to evade RNA silencing in addition to encoding silencing suppressor activities. Our results should have important implications in further studies on RNA-based mechanisms of host-pathogen interactions and the biological constraints that shape the evolution of infectious RNA structures.
Figures

Similar articles
-
Double-stranded RNA interferes in a sequence-specific manner with the infection of representative members of the two viroid families.Virology. 2008 Feb 5;371(1):44-53. doi: 10.1016/j.virol.2007.09.031. Epub 2007 Oct 29. Virology. 2008. PMID: 18028975
-
Comprehensive Transcriptome Analyses Reveal that Potato Spindle Tuber Viroid Triggers Genome-Wide Changes in Alternative Splicing, Inducible trans-Acting Activity of Phased Secondary Small Interfering RNAs, and Immune Responses.J Virol. 2017 May 12;91(11):e00247-17. doi: 10.1128/JVI.00247-17. Print 2017 Jun 1. J Virol. 2017. PMID: 28331096 Free PMC article.
-
Specific argonautes selectively bind small RNAs derived from potato spindle tuber viroid and attenuate viroid accumulation in vivo.J Virol. 2014 Oct;88(20):11933-45. doi: 10.1128/JVI.01404-14. Epub 2014 Aug 6. J Virol. 2014. PMID: 25100851 Free PMC article.
-
Viroid pathogenesis: a critical appraisal of the role of RNA silencing in triggering the initial molecular lesion.FEMS Microbiol Rev. 2020 May 1;44(3):386-398. doi: 10.1093/femsre/fuaa011. FEMS Microbiol Rev. 2020. PMID: 32379313 Review.
-
Potato Spindle Tuber Viroid RNA-Templated Transcription: Factors and Regulation.Viruses. 2018 Sep 17;10(9):503. doi: 10.3390/v10090503. Viruses. 2018. PMID: 30227597 Free PMC article. Review.
Cited by
-
A nuclear-replicating viroid antagonizes infectivity and accumulation of a geminivirus by upregulating methylation-related genes and inducing hypermethylation of viral DNA.Sci Rep. 2016 Oct 14;6:35101. doi: 10.1038/srep35101. Sci Rep. 2016. PMID: 27739453 Free PMC article.
-
Modification of tobacco plant development by sense and antisense expression of the tomato viroid-induced AGC VIIIa protein kinase PKV suggests involvement in gibberellin signaling.BMC Plant Biol. 2009 Aug 18;9:108. doi: 10.1186/1471-2229-9-108. BMC Plant Biol. 2009. PMID: 19689802 Free PMC article.
-
Appearances can be deceptive: revealing a hidden viral infection with deep sequencing in a plant quarantine context.PLoS One. 2014 Jul 25;9(7):e102945. doi: 10.1371/journal.pone.0102945. eCollection 2014. PLoS One. 2014. PMID: 25061967 Free PMC article.
-
First Detection and Molecular Characterization of Peach Latent Mosaic Viroid (PLMVd) in Kazakhstan.Pathogens. 2025 Mar 3;14(3):243. doi: 10.3390/pathogens14030243. Pathogens. 2025. PMID: 40137728 Free PMC article.
-
Application of RNA silencing to plant disease resistance.Silence. 2012 May 31;3(1):5. doi: 10.1186/1758-907X-3-5. Silence. 2012. PMID: 22650989 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
