

An essential role for IL-17 in preventing pathogen-initiated bone destruction: recruitment of neutrophils to inflamed bone requires IL-17 receptor-dependent signals
- PMID: 17202320
- PMCID: PMC1874584
- DOI: 10.1182/blood-2005-09-010116
An essential role for IL-17 in preventing pathogen-initiated bone destruction: recruitment of neutrophils to inflamed bone requires IL-17 receptor-dependent signals
Abstract
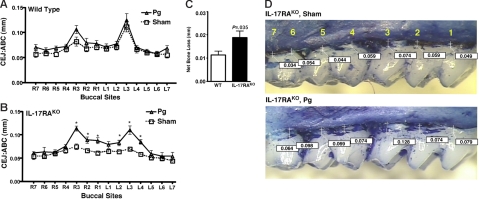
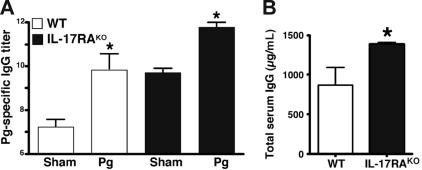
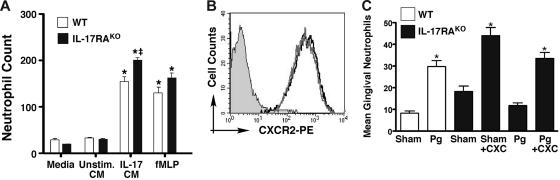
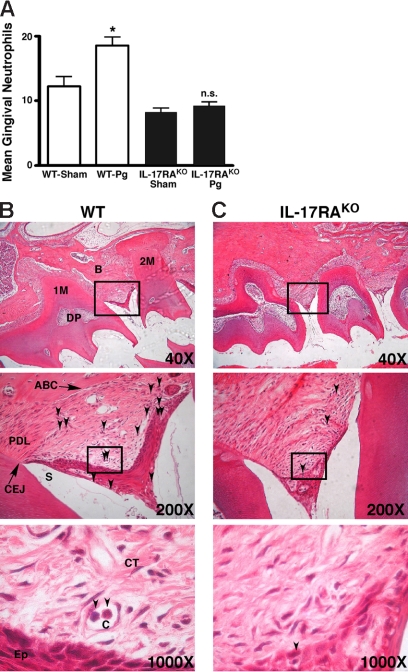
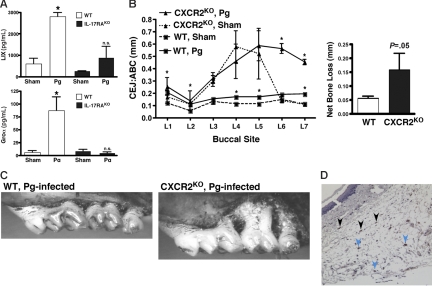
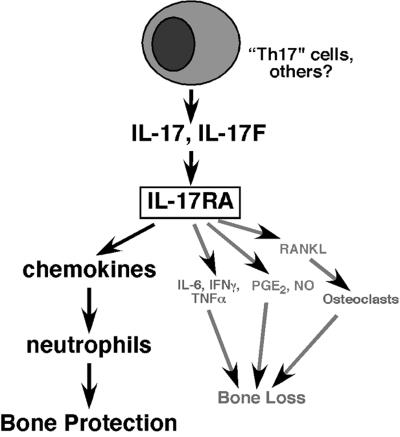
IL-17 and its receptor are founding members of a novel family of inflammatory cytokines. IL-17 plays a pathogenic role in rheumatoid arthritis (RA)-associated bone destruction. However, IL-17 is also an important regulator of host defense through granulopoiesis and neutrophil trafficking. Therefore, the role of IL-17 in pathogen-initiated bone loss was not obvious. The most common form of infection-induced bone destruction occurs in periodontal disease (PD). In addition to causing significant morbidity, PD is a risk factor for atherosclerotic heart disease and chronic obstructive pulmonary disease (COPD). Similar to RA, bone destruction in PD is caused by the immune response. However, neutrophils provide critical antimicrobial defense against periodontal organisms. Since IL-17 is bone destructive in RA but a key regulator of neutrophils, we examined its role in inflammatory bone loss induced by the oral pathogen Porphyromonas gingivalis in IL-17RA-deficient mice. These mice showed enhanced periodontal bone destruction, suggesting a bone-protective role for IL-17, reminiscent of a neutrophil deficiency. Although IL-17RA-deficient neutrophils functioned normally ex vivo, IL-17RA knock-out (IL-17RA(KO)) mice exhibited reduced serum chemokine levels and concomitantly reduced neutrophil migration to bone. Consistently, CXCR2(KO) mice were highly susceptible to alveolar bone loss; interestingly, these mice also suggested a role for chemokines in maintaining normal bone homeostasis. These results indicate a nonredundant role for IL-17 in mediating host defense via neutrophil mobilization.
Figures
References
-
- Moseley TA, Haudenschild DR, Rose L, Reddi AH. Interleukin-17 family and IL-17 receptors. Cytokine Growth Factor Rev. 2003;14:155–174. - PubMed
-
- Huang W, Na L, Fidel PL, Schwarzenberger P. Requirement of interleukin-17A for systemic anti-Candida albicans host defense in mice. J Infect Dis. 2004;190:624–631. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
