

Nonvesicular release of glutamate by glial xCT transporters suppresses glutamate receptor clustering in vivo
- PMID: 17202478
- PMCID: PMC2193629
- DOI: 10.1523/JNEUROSCI.4770-06.2007
Nonvesicular release of glutamate by glial xCT transporters suppresses glutamate receptor clustering in vivo
Abstract
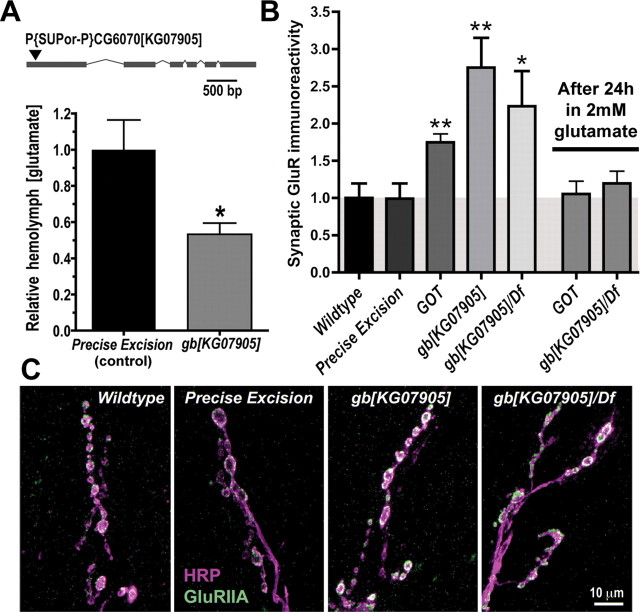
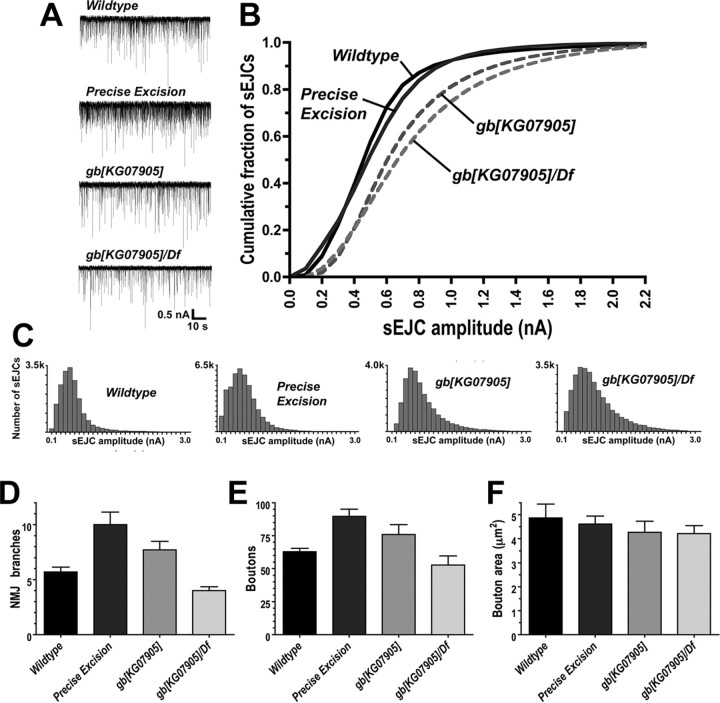
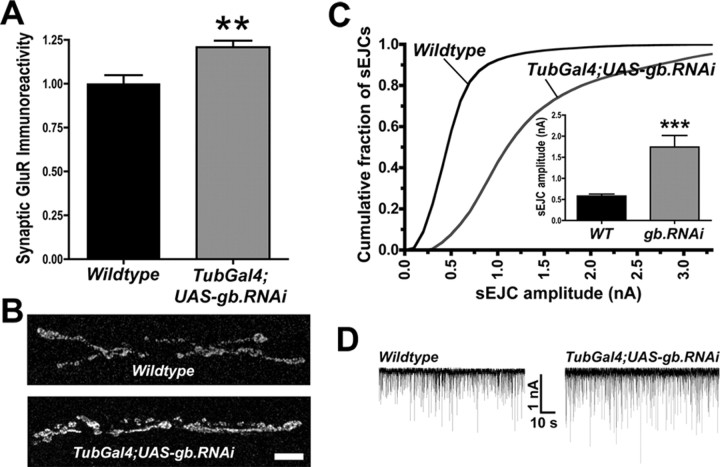
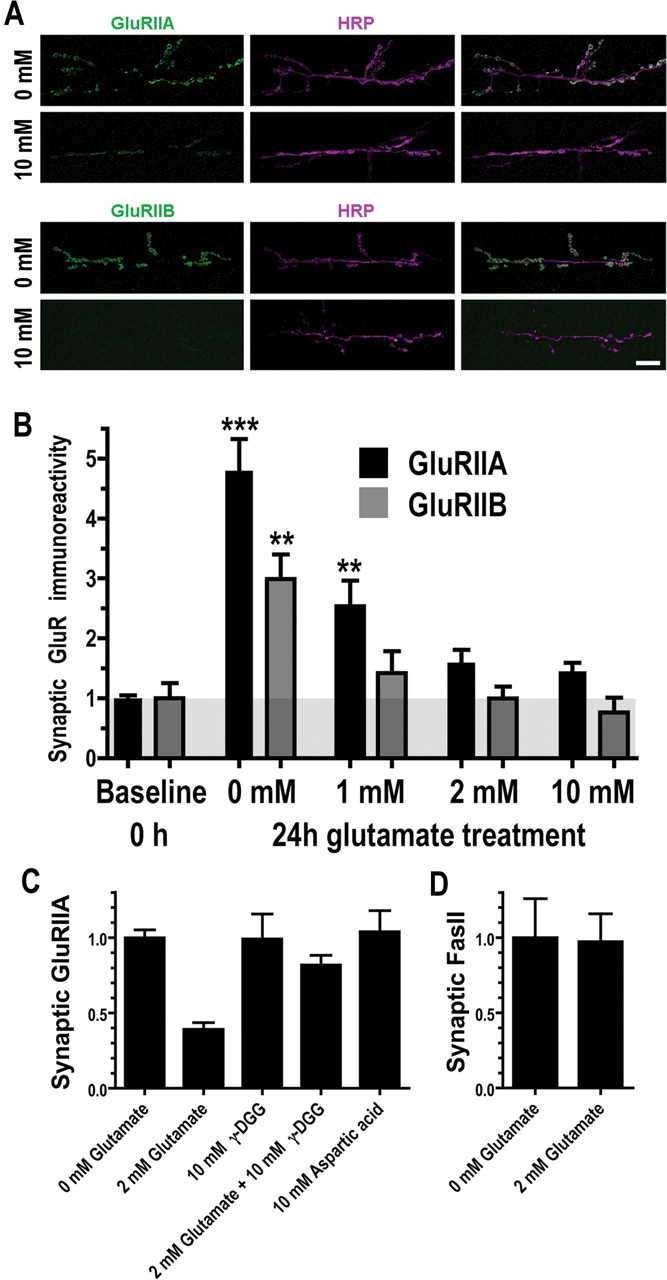
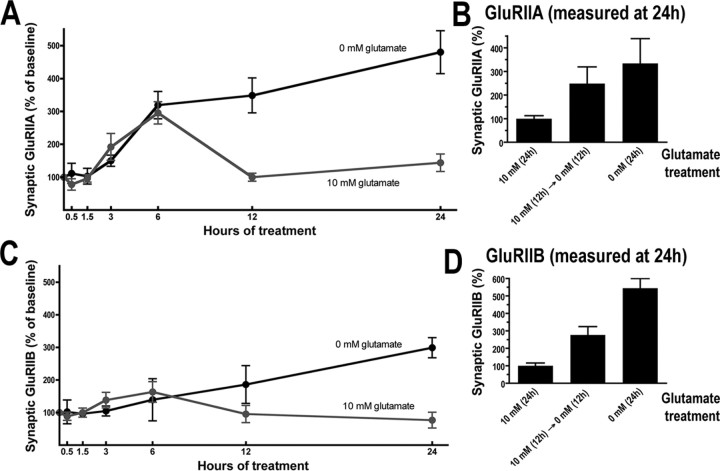
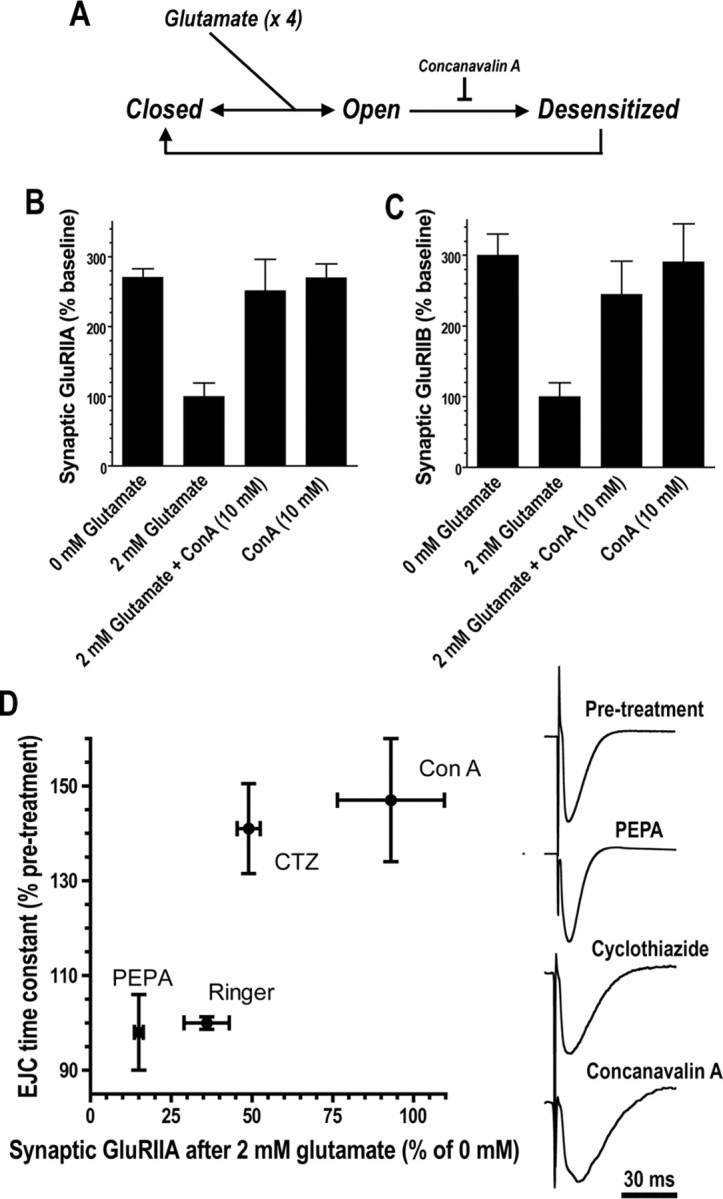
We hypothesized that cystine/glutamate transporters (xCTs) might be critical regulators of ambient extracellular glutamate levels in the nervous system and that misregulation of this glutamate pool might have important neurophysiological and/or behavioral consequences. To test this idea, we identified and functionally characterized a novel Drosophila xCT gene, which we subsequently named "genderblind" (gb). Genderblind is expressed in a previously overlooked subset of peripheral and central glia. Genetic elimination of gb causes a 50% reduction in extracellular glutamate concentration, demonstrating that xCT transporters are important regulators of extracellular glutamate. Consistent with previous studies showing that extracellular glutamate regulates postsynaptic glutamate receptor clustering, gb mutants show a large (200-300%) increase in the number of postsynaptic glutamate receptors. This increase in postsynaptic receptor abundance is not accompanied by other obvious synaptic changes and is completely rescued when synapses are cultured in wild-type levels of glutamate. Additional in situ pharmacology suggests that glutamate-mediated suppression of glutamate receptor clustering depends on receptor desensitization. Together, our results suggest that (1) xCT transporters are critical for regulation of ambient extracellular glutamate in vivo; (2) ambient extracellular glutamate maintains some receptors constitutively desensitized in vivo; and (3) constitutive desensitization of ionotropic glutamate receptors suppresses their ability to cluster at synapses.
Figures

References
-
- Aldred S, Moore KM, Fitzgerald M, Waring RH. Plasma amino acid levels in children with autism and their families. J Autism Dev Disord. 2003;33:93–97. - PubMed
-
- Armstrong N, Jasti J, Beich-Frandsen M, Gouaux E. Measurement of conformational changes accompanying desensitization in an ionotropic glutamate receptor. Cell. 2006;127:85–97. - PubMed
-
- Ball R, Xing B, Bonner P, Shearer J, Cooper RL. Long-term in vitro maintenance of neuromuscular junction activity of Drosophila larvae. Comp Biochem Physiol A Mol Integr Physiol. 2003;134:247–255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases