

A natural genetic code expansion cassette enables transmissible biosynthesis and genetic encoding of pyrrolysine
- PMID: 17204561
- PMCID: PMC1783357
- DOI: 10.1073/pnas.0610294104
A natural genetic code expansion cassette enables transmissible biosynthesis and genetic encoding of pyrrolysine
Abstract
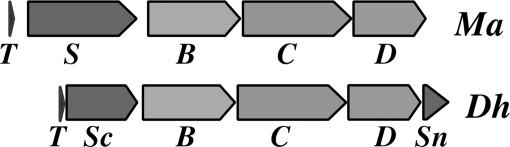
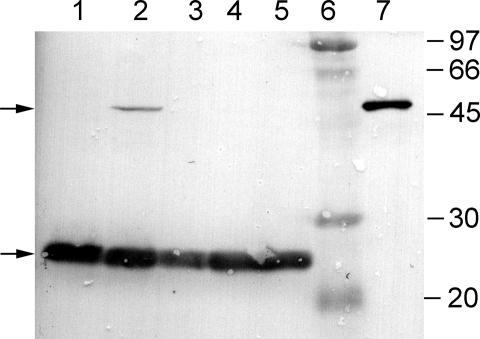
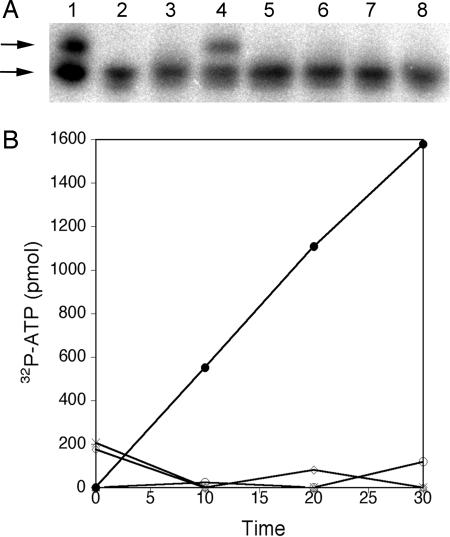
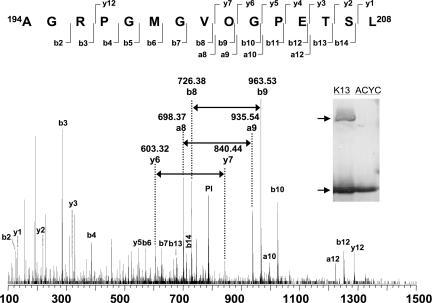
Pyrrolysine has entered natural genetic codes by the translation of UAG, a canonical stop codon. UAG translation as pyrrolysine requires the pylT gene product, an amber-decoding tRNA(Pyl) that is aminoacylated with pyrrolysine by the pyrrolysyl-tRNA synthetase produced from the pylS gene. The pylTS genes form a gene cluster with pylBCD, whose functions have not been investigated. The pylTSBCD gene order is maintained not only in methanogenic Archaea but also in a distantly related Gram-positive Bacterium, indicating past horizontal gene transfer of all five genes. Here we show that lateral transfer of pylTSBCD introduces biosynthesis and genetic encoding of pyrrolysine into a naïve organism. PylS-based assays demonstrated that pyrrolysine was biosynthesized in Escherichia coli expressing pylBCD from Methanosarcina acetivorans. Production of pyrrolysine did not require tRNA(Pyl) or PylS. However, when pylTSBCD were coexpressed with mtmB1, encoding the methanogen monomethylamine methyltransferase, UAG was translated as pyrrolysine to produce recombinant monomethylamine methyltransferase. Expression of pylTSBCD also suppressed an amber codon introduced into the E. coli uidA gene. Strains lacking one of the pylBCD genes did not produce pyrrolysine or translate UAG as pyrrolysine. These results indicated that pylBCD gene products biosynthesize pyrrolysine using metabolites common to Bacteria and Archaea and, furthermore, that the pyl gene cluster represents a "genetic code expansion cassette," previously unprecedented in natural organisms, whose transfer allows an existing codon to be translated as a novel endogenously synthesized free amino acid. Analogous cassettes may have served similar functions for other amino acids during the evolutionary expansion of the canonical genetic code.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- James CM, Ferguson TK, Leykam JF, Krzycki JA. J Biol Chem. 2001;276:34252–34258. - PubMed
-
- Hao B, Gong W, Ferguson TK, James CM, Krzycki JA, Chan MK. Science. 2002;296:1462–1466. - PubMed
-
- Hao B, Zhao G, Kang P, Soares J, Ferguson T, Gallucci J, Krzycki J, Chan M. Chem Biol. 2004;11:1317–1324. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
